10. Гаметогенез и развитие растений. Размножение организмов
10. Гаметогенез и развитие растений
Мейоз в жизненном цикле растений. У растений гаметогенез и размножение протекают иначе, чем у животных. Процесс мейоза происходит у них не на стадии образования гамет, а на стадии образования спор. Кроме того, у растений наблюдается чередование поколений с диплоидным (2n) и гаплоидным (n) набором хромосом.
Поколение с гаплоидным набором хромосом называется гаметофитом. На гаметофите образуются гаметы в процессе митоза. Поколение с диплоидным набором хромосом называется спорофитом, и на нем образуются споры в процессе мейоза. Гаметофит развивается из гаплоидных спор, а спорофит — из диплоидной зиготы, образующейся в результате оплодотворения.
Смена поколений идет по схеме: зигота (2n) —> спорофит (2n) —> мейоз —>
В цикле развития мейоз всегда происходит один раз. В зависимости от периода жизни спорофита и гаметофита взрослое растение может быть гаплоидным или диплоидным.
Размножение и развитие зеленых водорослей
У низших растений преобладающим поколением является гаметофит. Он размножается бесполым путем, образуя клетки, из которых развиваются взрослые особи. В определенный период на гаметофите образуются гаметы, разные или одинаковые по величине.
После слияния гамет образуется зигота, которая сразу же делится мейозом и образует споры, дающие начало новым гаметофитам. Таким образом, в жизненном цикле зеленых водорослей диплоидное поколение представлено только одной клеткой — зиготой.
Размножение и развитие высших споровых растений
У мхов, папоротников, плаунов и хвощей размножение происходит спорами.
У мхов взрослым растением является гаметофит (n) — половое поколение, которое развивается при прорастании споры (рис. 19).
Рис. 19. Схема образования (А) и развития (Б) зародышевого мешка: 1 — антиподы; 2 — два центральных ядра; 3 — синергиды; 4 — яйцеклетка

Это листостебельное растение, на побегах которого развиваются органы полового размножения — антеридии (мужские органы) и архегонии (женские органы). В антеридиях в процессе митоза образуются сперматозоиды, а в архегониях — яйцеклетка (как правило, одна). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку, при этом образуется зигота (2n). Из зиготы на гаметофите развивается спорофит (2n) в виде коробочки на ножке — спорогона. Спорофит не способен к самостоятельному существованию и питается за счет питательных веществ гаметофита. В спорангиях (коробочке) в результате мейоза образуются споры. Споры после созревания высыпаются и во влажной среде прорастают, давая начало новым гаметофитам.
Схема жизненного цикла мхов

У папоротников, плаунов и хвощей, наоборот, взрослым растением является спорофит, на котором в специальных органах — спорангиях в результате мейоза образуются споры (n). Споры после созревания высыпаются и прорастают (рис. 20).
Рис. 20. Схема образования (А) и развития (Б) пыльцевого зерна: 1 — вегетативная клетка; 2 — генеративная клетка

При прорастании споры развивается половое поколение — гаметофит, который представлен небольшим заростком и существует очень недолго. На заростке развиваются органы полового размножения — антеридии и архегонии, в которых в результате митоза развиваются гаметы. В антеридиях образуются сперматозоиды, а в архегониях созревает яйцеклетка.
Схема жизненного цикла папоротников

При наличии капельно-жидкой влаги сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку, при этом образуется зигота. Из зиготы развивается зародыш, а далее молодое растение — спорофит.
Размножение и развитие семенных растений
У семенных растений размножение происходит семенами. Преобладающим поколением является спорофит, а гаметофит сильно редуцирован, развивается в спорофите и представлен лишь несколькими клетками.
Схема жизненного цикла голосеменных (на примере сосны)

Процесс развития семенных растений рассмотрим более подробно на примере цветковых растений. Взрослое растение имеет диплоидный набор хромосом и является спорофитом. Оно развивается из семени.
Репродуктивным органом является цветок. В цветке образуются женский орган — пестик и мужской — тычинки.
В завязи пестика в семязачатках из спорогенной ткани в результате мейоза образуются 4 споры (n), причем одна из них, крупная — мегаспора развивается в женский заросток — гаметофит; три другие, более мелкие, отмирают.
Мегаспора трижды делится митозом, и образуется восьмиядерный зародышевый мешок. Восемь ядер распределяются следующим образом. Ближнее к пыльцевходу крупное ядро — яйцеклетка, рядом два ядра помельче — две сопутствующие клетки — синергиды. На противоположном полюсе мешка располагаются три ядра — антиподы, а в центре располагаются два центральных ядра. Все ядра гаплоидные. Таким образом, женский гаметофит представлен восьмиядерным зародышевым мешком.
В тычинках, в пыльцевых мешках из ткани микроспорангия в результате мейоза образуется много мелких микроспор (n). Все споры развиваются и дают начало мужскому заростку — гаметофиту. Спора делится митозом и образует вегетативную и генеративную клетки. Ядро генеративной клетки делится еще раз, и образуется два спермия. Вегетативная и генеративная клетки покрываются оболочкой, образуется пыльцевое зерно. Таким образом, мужской гаметофит представлен двумя клетками.
При попадании пыльцы на рыльце пестика вегетативная клетка начинает прорастать, образуя пыльцевую трубку и продвигая генеративную клетку к пыльцевходу. Два спермия через пыльцевход проникают в зародышевый мешок. Один спермий сливается с яйцеклеткой, и образуется зигота (2n), из которой развивается зародыш семени. Другой спермий сливается с двумя ядрами центральной клетки, в результате чего образуется эндосперм (3n) семени, в котором запасаются питательные вещества.
Схема жизненного цикла покрытосеменных растений

Этот процесс называется двойным оплодотворением. Он был открыт русским ученым С. П. Навашиным. В результате двойного оплодотворения в семязачатке образуется семя, а из покрова семязачатка — семенная кожура. Вокруг семени из завязи и других частей цветка образуется плод (рис. 21).
Рис. 21. Схема двойного оплодотворения цветковых растений: 1 — пыльца; 2 — пыльцевая трубка вегетативной клетки; 3 — два спермия; 4 — яйцеклетка; 5 — два центральных ядра; 6 — семязачаток; 7 — эндосперм семени; 8 — зародыш; 9 — кожура семени из покрова семязачатка

У растений, при переходе от низших к высшим, наблюдается постепенное увеличение срока жизни спорофита. Начиная с папоротникообразных, у всех растений взрослый организм представлен спорофитом, а гаметофит претерпевает постепенно редукцию до одной или нескольких клеток.
Вопросы для самоконтроля
1. В чем заключается особенность жизненного цикла растений по сравнению с животными?
2. Как происходит чередование поколений у растений?
3. Что такое гаметофит и спорофит? В чем их отличие?
4. Какое поколение является господствующим у водорослей и мхов, папоротникообразных и семенных растений?
5. Как изменяется гаметофит от низших растений к высшим?
6. В чем их сходство и отличие гаметофита цветковых растений и мхов?
7. Какой процесс называется двойным оплодотворением? В чем его особенность?
Поделитесь на страничкеСледующая глава >
bio.wikireading.ru
Биология для студентов — 09. Гаметогенез у растений
У растений процесс формирования половых клеток подразделяется на два этапа:
- 1-й этап—спорогенез, который завершается образованием вегетативных гаплоидных спор,
- 2-й этап—собственно гаметогенез, в течение которого образуются зрелые гаметы.
Процесс образования мужских половых клеток складывается из:
- микроспорогенеза (образования микроспор),
- микрогаметогенеза—созревания спермиев внутри пыльцевого зерна.
Процесс образования женских половых клеток складывается из:
- мегаспорогенеза (или макро-спорогенеза),
- мегагаметогенеза—формирования зрелого зародышевого мешка, в котором образуется яйцеклетка

Процесс микроспорогенеза у цветковых растений протекает в пыльниках, причем еще в то время, когда бутон не раскрылся или, когда колос у злаковых еще в трубке, у многолетних растений все это происходит осенью. В основе микроспорогенеза лежит мейоз, в результате которого образуются четыре споры (гаплоидные вегетативные клетки). Образование пыльцевого зерна (микрогаметогенез) начинается с митотического деления ядра споры. Ядро делится митозом на два, и образуются две клетки—вегетативная и генеративная. Вегетативная клетка обладает запасом питательных веществ, которые необходимы для обеспечения роста пыльцевой трубки при прорастании пыльцы на рыльце пестика и для деления генеративной клетки на две, которые и представляют собой собственно мужские половые клетки растений. Они не обладают способностью к самостоятельному движению и называются
Мегаспорогенез протекает в семяпочке цветковых растении, когда одна из клеток субэпидермального слоя (археспориальная клетка) обособляется, растет и превращается в материнскую клетку мегаспоры.

Этот процесс протекает во время цветения растений. В материнской клетке происходит мейоз, в результате которого образуются четыре клетки—мегаспоры. У большинства высших растений только одна развивается дальше, остальные три гибнут (моноспорический тип развития).
После образования мегаспоры, имеющей одно гаплоидное ядро, начинается процесс формирования зародышевого мешка, в котором и образуется собственно яйцеклетка, т. е. начинается процесс мегагаметогенеза. Мегаспора растет, и у большинства растений ядро ее претерпевает три митотических деления. Крупная клетка с восьмью одинаковыми ядрами называется зародышевым мешком. Вокруг ядер обособляется плазма, поэтому эти образования иногда называют зародышевыми клетками. Две клетки, находящиеся у микропиле—входа в зародышевый мешок, называются синергидами. Они играют вспомогательную роль при оплодотворении и скоро погибают. Одна клетка—это собственно яйцеклетка. Две клетки называются полярными, при перемещении их в центр зародышевого мешка они сливаются, образуя одно диплоидное ядро. Оно называется центральным ядром зародышевого мешка. Последние три клетки чаще всего находятся на стороне, противоположной микропиле, и называются антиподами. Они играют вспомогательную роль в начале развития зиготы и затем погибают.
vseobiology.ru
Биология для студентов — 50. Спорогенез и гаметогенез у растений. Общая схема жизненного цикла растений
Процесс формирования половых клеток у растений подразделяется на два этапа:
- спорогенез — завершается образованием гаплоидных клеток — спор,
- гаметогенез — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки — это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит — это зародышевый мешок, который закладывается и развивается внутри семяпочки.

Схема мегаспорогенеза (1-5) и мегагаметогенеза (6-12) у растений
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры. В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других — в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения. Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно. В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других — из двух (биспорический тип) и четырех спор (тетраспорический тип).
В ходе жизненного цикла растений происходит смена двух типов организмов: поколение гаплоидного гаметофита и поколение диплоидного спорофита поочередно сменяют друг друга, что схематически показано на рисунке.

Гаплидное поколение называется гаметофитом (от греч. gamete — жена, gametes — муж; phyton — растение), так как оно способно к половому размножению и образует гаметы. Поскольку образование гамет происходит в результате митоза, они тоже гаплоидны. Сливаясь, гаметы образуют диплоидную зиготу, из которой вырастает следующее поколение — поколение диплоидных спорофитов. Они называются спорофитами, потому что способны к бесполому размножению с образованием спор. Споры образуются в результате мейоза, т. е. здесь происходит возврат к гаплоидному состоянию. Гаплоидные споры дают начало гаметофитному поколению. Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла; это поколение называют доминантным. У моховидных доминирует поколение гаметофитов, у всех остальных наземных растений — поколение спорофитов. Доминирующее поколение принято помещать в верхнюю половину схемы, изображающей жизненный цикл.
vseobiology.ru
Спорогенез и гаметогенез у растений
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или
www.activestudy.info
1. Спорогенез и гаметогенез у растений. Спорогенез и гаметогенез у растений
Похожие главы из других работ:
Вегетативное размножение хвойных растений
2. Характеристика маточных растений
2.1 Туя западная — Thuja occidentalis В природе встречается в восточных районах Северной Америки. Однодомное дерево 12-20 м высотой, реже кустарник. Крона компактная, в молодости узкопирамидальная и яйцевидная в зрелом возрасте…
Вирусы и их особенности
2.1 Вирусы растений
О том, что растения болеют, люди узнали в те далекие времена, когда перешли на оседлое земледелие. Земледельцы как могли, лечили растения, старались предотвратить массовое поражение…
Закаливание растений
1. Закаливание растений
…
Закаливание растений
2. Холодостойкость растений
Устойчивость растений к низким температурам подразделяют на холодостойкость и морозоустойчивость. Под холодостойкостью понимают способность растений переносить положительные температуры несколько выше О 0С…
Закаливание растений
3. Морозоустойчивость растений
Морозоустойчивость — способность растений переносить температуру ниже О °С, низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур…
Клумбы и цветочные растения
2.1 История клумбовых растений
Некоторые считают, что в Британии высаживать цветы на клумбы впервые стали только во времена королевы Виктории, когда в моде было все яркое. Однако в действительности история клумбовых растений началась задолго до того, как в 1837 г…
Клумбы и цветочные растения
2.2 Типы клумбовых растений
Теплолюбивые однолетники Жизненный цикл теплолюбивых однолетников длится один сезон: их выращивают из семян, летом или осенью они цветут и затем отмирают. Они не переносят низких температур, поэтому для того…
Концепции современного естествознания
3.4 Селекция растений и животных
Одним из самых распространенных достижений человеческой цивилизации было выведение сортов растений и пород домашних животных от диких предков. Отбирая те особи, которые обладали какими-то желательными отклонениями…
Методы селекции
Методы селекции растений
Основные методы селекции растений в частности — отбор и гибридизация. Для перекрестно-опыляемых растений применяют массовый отбор особей с желаемыми свойствами. В противном случае невозможно получить материал для дальнейшего скрещивания…
Пластиды и их пигменты. Выделительные системы растений
VI. ПОНЯТИЕ О ВИДЕ РАСТЕНИЙ. СОСТАВЛЕНИЕ ВИДОВЫХ НАЗВАНИЙ РАСТЕНИЙ СОГЛАСНО БИНАРНОМУ МЕТОДУ К. ЛИННЕЯ. ВЫПИШЕТЕ ИЗ «СПИСКА ОСНОВНЫХ СЕМЕЙСТВ И ВИДОВ» 6 ВИДОВ (ИЗ НИХ 2 ОДНОГО РОДА) ИЗ РАЗНЫХ СЕМЕЙСТВ
Вид — совокупность популяций особей, способных к скрещиванию с образованием плодовитого потомства, населяющих определенную территорию…
Процесс и проблемы клонирования
4.1 Клонирование растений
Клонирование растений, в отличие от клонирования животных, является обычным процессом, с которым сталкивается любой цветовод или садовод. Ведь часто растение размножают отростками, черенками, усиками и т.д. Это и есть пример клонирования…
Транспирация растений
1.4 Значение транспирации для растений
Основное значение транспирации заключается в охлаждении растений. Поглощая солнечный лучи в процессе фотосинтеза, растение, естественно, сильно нагреваются…
Ультрафиолет как стресс-фактор для растений
2.5 Понятие стресса растений
Способность к защите от повреждающих и неблагоприятных факторов среды — обязательное свойство любого, в том числе и растительного организма. Ответные реакции, индуцируемые в организме внешними воздействиями…
Этапы гаметогенеза и виды скрещивания
2. Половое размножение. Развитие половых клеток (гаметогенез)
У более сложно организованных животных или исключительно, или, по крайней мере, преимущественно, практикуется половой способ размножения, хоть зачатки этой формы воспроизведения потомства, так сказать намек на него…
Яды животных и растений
2. Яды растений
Яды растений также можно разделить на белковые и небелковые. Выделенные и охарактеризованные яды белковой природы относительно немногочисленны. Так, в бледной поганке и некоторых мухоморах содержатся фаллотоксины и аматотоксины…
bio.bobrodobro.ru
14.Спорогенез, гаметогенез и оплодотворение у покрытосеменных растений.
Спорогенез, гаметогенез и оплодотворение у растений.
У Покрытосеменных растений спорогенез, гаметогенез и оплодотворение представляют собой непрерывную последовательность репродуктивных процессов, происходящих в цветках. В типичных цветках репродуктивные органы представлены гинецеем (совокупностью плодолистиков, образующих женскую репродуктивную сферу) и андроцеем (совокупностью тычинок, образующих мужскую репродуктивную сферу).
Мегаспорогенез, образование зародышевого мешка и яйцеклетки
Мегаспорогенез протекает в женской репродуктивной сфере – в гинецее. Морфологически гинецей представлен пестиком (или несколькими пестиками). В состав отдельного пестика входят: завязь, столбик и рыльце. Внутри завязи находятся семязачатки (один или несколько). Внутреннее содержимое семязачатка называется нуцеллус; снаружи семязачаток окружен покровами (одиночным или двойным интегументом). В нуцеллусе семязачатка (2n) содержится одна археспориальная клетка, способная делиться путем мейоза (у некоторых растений археспорий многоклеточный). В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре мегаспоры. Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит). Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра – одно центральное диплоидное ядро; два ядра – две клетки-синергиды; одно ядро становится ядром яйцеклетки.
Микроспорогенез и образование пыльцевых зерен
Микроспорогенез протекает в мужской репродуктивной сфере – в андроцее. Каждый элемент андроцея – тычинка – состоит из тычиночной нити и пыльника. Внутри пыльника имеются пыльцевые гнезда, содержащие археспориальную ткань. В археспориальной ткани пыльников из каждой диплоидной материнской клетки микроспор в результате мейоза образуется четыре микроспоры. Каждая микроспора делится путем митоза и образуется двухклеточное пыльцевое зерно: одна клетка называется генеративной (в дальнейшем на ее основе образуются спермии), вторая – вегетативной (это остаток вегетативных клеток мужского гаметофита). Зрелое пыльцевое зерно покрыто двойной оболочкой: экзиной (внешней) и интиной (внутренней).
Опыление
Опыление – это процесс переноса пыльцы из пыльников на рыльце пестика. Существует два основных типа опыления: самоопыление и перекрестное опыление. При самоопылении происходит перенос пыльцы в пределах одного цветка, а при перекрестном опылении пыльца из пыльников одного цветка переносится на рыльце пестика другого цветка.
Для более точной характеристики опыления с точки зрения генетики используются специальные термины: клейстогамия (самоопыление в нераскрывшемся цветке), автогамия (самоопыление в раскрывшемся цветке), гейтоногамия (перенос пыльцы из пыльника одного цветка на рыльце пестика другого цветка в пределах одного растения), аллогамия (перенос пыльцы из пыльника цветка одного растения на рыльце пестика цветка другого растения). Таким образом, лишь аллогамия является собственно перекрестным опылением, а остальные типы опыления тождественны самоопылению.
Образование пыльцевой трубки и спермиев
В результате опыления пыльцевое зерно попадает на рыльце пестика и прорастает в пыльцевую трубку (мужской гаметофит). В пыльцевой трубке ядро генеративной клетки делится путем митоза, образуя два генеративных ядра. (У ряда растений деление генеративной клетки происходит еще в пыльниках). Каждое генеративное ядро с прилежащим слоем цитоплазмы называется спермием.
Двойное оплодотворение, образование зародыша и семени
Пыльцевая трубка внедряется в ткани столбика и растет в направлении завязи, при этом на вершине пыльцевой трубки находится вегетативное ядро, а за ним движутся оба спермия. Затем пыльцевая трубка проникает в семязачаток через пыльцевход – микропиле (у некоторых растений существует халазогамия – проникновение пыльцевой трубки в семязачаток через его ножку – халазу). Далее пыльцевая трубка лопается, ее содержимое изливается в зародышевый мешок, и происходит двойное оплодотворение. Один спермий сливается с ядром яйцеклетки, и образуется диплоидная зигота. Другой спермий сливается с центральным диплоидным ядром зародышевого мешка, и образуется триплоидное ядро эндосперма. Все остальные ядра зародышевого мешка и вегетативное ядро пыльцевого зерна – разрушаются.
Из диплоидной зиготы развивается зародыш, а из триплоидного центрального ядра – эндосперм. Происходит формирование семени, включающего зародыш, эндосперм и семенную кожуру, которая образуется из покровов семязачатка.
studfile.net
3.3.2. Гаметогенез
Половые клетки (гаметы) отличаются от соматических клеток: 1) гаплоидным набором хромосом; 2) резко увеличенными (яйцеклетка) или резко уменьшенными (сперматозоид) размерами; это связано с тем, что яйцеклетка накапливает питательные вещества (желток) для развивающегося из зиготы зародыша, а сперматозоид лишь перемещает наследственный материал (гаплоидный набор хромосом) к яйцеклетке; 3) низким уровнем обменных процессов, напоминающим таковой при состоянии анабиоза.
Процесс развития и образования половых клеток называется гаметогенезом. Существенным этапом в этом процессе является мейоз, однако он не исчерпывает всего процесса гаметогенеза, который обладает спецификой у особей разных полов и у представителей разных групп организмов.
У животных, в отличие от растений, в онтогенезе очень рано обособляются зачатковые клетки, которые впоследствии дают начало половым железам и половым клеткам. Зачатковые клетки делятся митозом и образуют гонии. Сначала они одинаковы у особей разных полов, затем дифференцируются у самцов в сперматогонии, у самок — в оогонии. Дальнейший процесс их формирования идет по-разному и носит название соответственно сперматогенеза и оогенеза (рис. 45).
В процессе сперматогенеза клетки проходят четыре периода (стадии): размножение, рост, созревание, формирование. Сперматогонии делятся митозом с сохранением диплоидного числа хромосом (период размножения). Затем деление прекращается, клетка растет и готовится к мейозу (период роста). В это время она имеет название сперматоцит I (первого) порядка. Сперматоцит I вступает в период созревания и претерпевает мейоз (рис. 45). В результате первого деления мейоза (у животных оно называется первым делением созревания и у большинства бывает редукционным) образуются две гаплоидные клетки, называемые сперматоцитами II (второго) порядка. Последние делятся еще раз (второе деление созревания, эквационное, или митотическое деление) и образуют сперматиды. Таким образом, из одного сперматогония образуются четыре сперматиды (рис. 45), которые вступают в фазу формирования сперматозоида, т.е. зрелой мужской гаметы (период формирования). Сперматозоиды животных имеют иногда очень сложное строение, необходимое для обеспечения им возможности слияния с женской половой клеткой (оплодотворение) в той или иной внешней среде. Сперматозоид состоит из головки, шейки и хвоста (рис. 46).
Сперматогенез у животных, в частности, у млекопитающих, начинается с момента закладки половых желез в эмбриогенезе. После рождения самца он прекращается и возобновляется вновь после полового созревания и протекает постоянно в течение всего периода зрелости. Этот процесс осуществляется в половых органах — семенниках. По выходу из семенника зрелые сперматозоиды приобретают в специальных органах (придатках) под влиянием многочисленных гормонов устойчивость к неблагоприятным факторам среды.
В оогенезе клетка проходит в основном те же периоды (стадии), что и в сперматогенезе, однако существует ряд особенностей (рис. 45). Так, после прекращения делений ооцит I (диплоидная клетка), в отличие

Рис. 45. Схема оогенеза и сперматогенеза
от сперматоцита I, проходит более выраженную стадию роста. В это время в цитоплазме ооцитов откладывается запас питательных веществ, необходимых для развития зародыша, вследствие чего ооциты увеличиваются в размерах.
П
Рис. 46. Строение сперматозоида:
1 – головка; 2 – шейка; 3 — хвост; 4 — акросома; 5 — ядро; 6 – центриоль; 7 — митохондрии
осле этого ооцитI, так же как и сперматоцит I, вступает в мейоз. В результате первого деления созревания образуются две гаплоидные клетки, которые, однако, резко отличаются друг от друга. Одна, крупная, сохраняющая цитоплазму и весь запас питательных веществ, называется ооцитом II. Другая, значительно меньшая клетка, является неполноценной и представляет собой выделившееся под оболочку первой клетки ядро. Её называют редукционным тельцем (полярным, или направительным тельцем, или же оотидой). Редукционное тельце дегенерирует. Иногда перед дегенерацией редукционное тельце успевает разделиться еще раз, образуя два редукционных тельца, судьба которых всегда одинакова: оба дегенерируют. Ооцит II делится (второе деление созревания) и образует снова две неравноценные клетки: одна — зрелая яйцеклетка, имеющая гаплоидный набор хромосом и несущая весь запас питательных веществ, другая клетка представляет собой второе направительное тельце. Таким образом, в отличие от сперматогенеза, из одного оогония образуется только одна функционирующая яйцеклетка. Яйцеклетка животных обычно округлой формы, лишена структурных усложнений, поэтому стадия формирования в оогенезе, по существу, не выражена (рис. 45).Оогенез у млекопитающих происходит в яичниках и тоже начинается в период эмбрионального развития. Так, у женского эмбриона человека в возрасте пяти месяцев половые клетки бывают уже на стадии ооцита I. После рождения оогенез вначале приостанавливается (на стадии ооцита I), а затем вновь продолжается с момента полового созревания: яйцеклетки развиваются из ооцитов I, возникших в эмбриогенезе. В течение всего периода половой зрелости оогенез протекает циклически (менструальные циклы у человека, астральные циклы у животных).
У растений процесс формирования половых клеток подразделяется на два этапа: 1) спорогенез, который завершается образованием вегетатив-
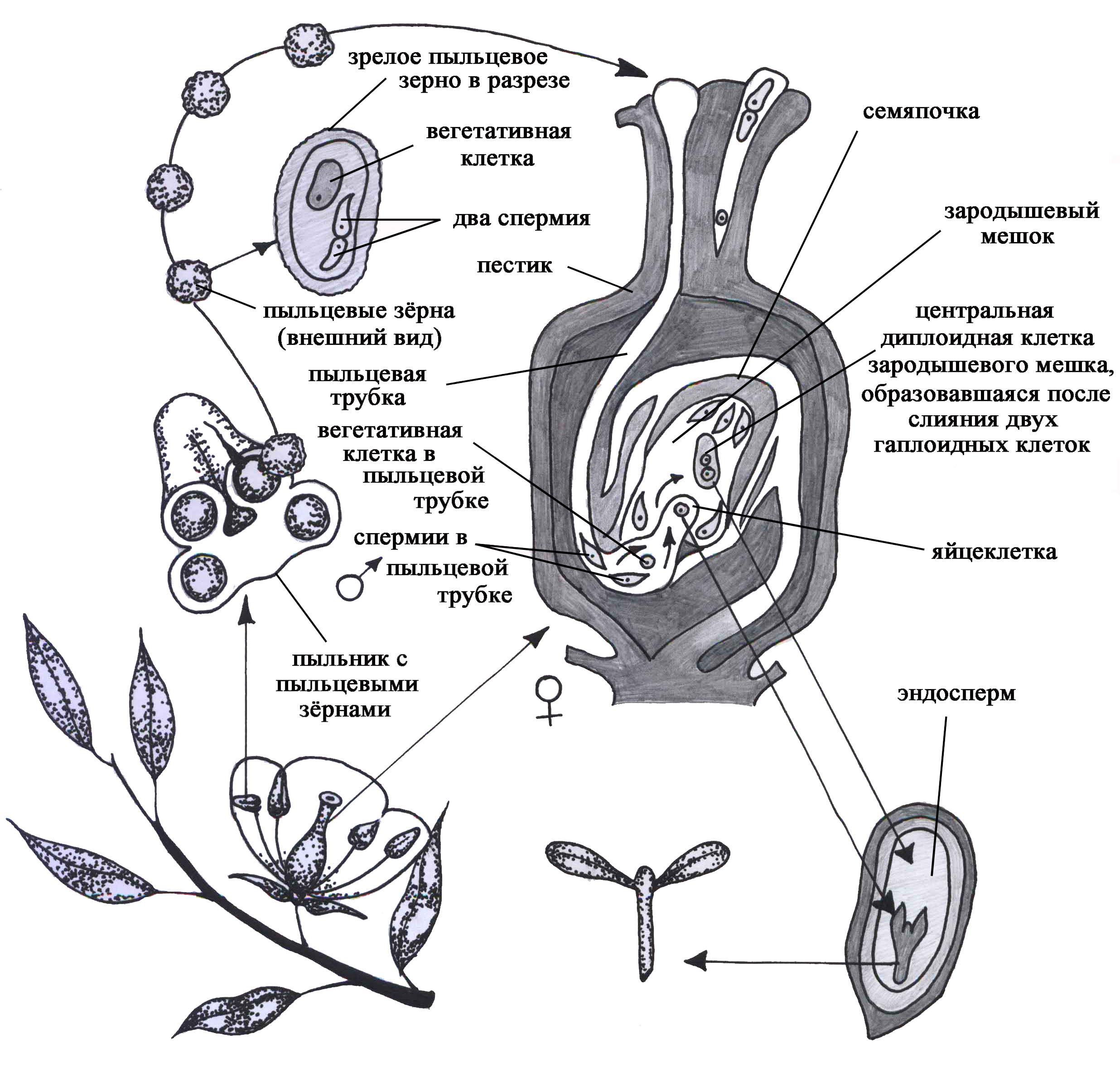
Рис. 47. Формирование половых клеток и оплодотворение у покрытосеменных растений
ных гаплоидных спор, 2) собственно гаметогенез, в течение которого образуются зрелые гаметы. Процесс образования мужских половых клеток складывается из микроспорогенеза (образования микроспор) и микрогаметогенеза — созревания спермиев внутри пыльцевого зерна. Образование женских половых клеток включает мегаспорогенез (или макроспорогенез) и мегагаметогенез — формирование зрелого зародышевого мешка, в котором образуется яйцеклетка.
Процесс микроспорогенеза у цветковых растений протекает в пыльниках (рис. 47), причем до раскрытия бутона или в период нахождения колоса злаковых еще в трубке. В основе микроспорогенеза лежит мейоз, в результате которого образуются четыре споры (гаплоидные вегетативные клетки). Образование пыльцевого зерна (микрогаметогенез) начинается с митотического деления ядра споры. В результате митотического деления ядра образуются вегетативная и генеративная клетки (рис. 47). Вегетативная клетка обладает запасом питательных веществ, которые необходимы: 1) для обеспечения роста пыльцевой трубки при прорастании пыльцы на рыльце пестика; 2) для деления генеративной клетки на две, которые и представляют собой собственно мужские половые клетки растений — спермии. Последние не обладают способностью к самостоятельному движению.
Мегаспорогенез протекает во время цветения в семяпочке цветковых растений (рис. 47). Материнская клетка вступает в два деления мейоза, в результате которых образуются четыре клетки — мегаспоры. У большинства высших растений только одна из них развивается дальше, а остальные три гибнут (моноспорический тип развития).
После образования мегаспоры, имеющей одно гаплоидное ядро, начинается процесс формирования зародышевого мешка, в котором и образуется собственно яйцеклетка, т.е. начинается процесс мегагаметогенеза. Мегаспора растет, и у большинства растений ее ядро претерпевает три митотических деления. Крупная клетка, содержащая 8 одинаковых ядер, называется зародышевым мешком. Вокруг ядер обособляется цитоплазма, поэтому эти образования иногда называют зародышевыми клетками. Две клетки, находящиеся у микропиле — входа в зародышевый мешок, называются синергидами. Они играют вспомогательную роль при оплодотворении и затем погибают. Одна клетка представлена соответственно яйцеклеткой. Две полярные клетки, перемещаясь в центр зародышевого мешка, сливаются, образуя центральную диплоидную клетку зародышевого мешка (рис. 47). Три остальные клетки (антиподы) чаще всего находятся на стороне, противоположной микропиле.
Сравнение процессов развития и созревания половых клеток у животных и растений указывает на их почти полный параллелизм, несмотря на то, что расхождение (дивергенция) растений и животных в филогенезе произошло на очень раннем этапе возникновения клеточной организации жизни. Это является одним из многих подтверждений однотипности принципов устройства ряда приспособительных механизмов в царствах растений и животных.
Зрелые сперматозоид и яйцо различаются своим строением, сходство свойственно только их ядрам. Тем не менее, обе гаметы образуются из одинаковых на вид первичных половых клеток. У всех организмов, размножающихся половым путем, эти первичные половые клетки обособляются на ранних стадиях развития от других клеток и развиваются особым образом, готовясь к выполнению своей функции — продуцированию половых, или зародышевых клеток. Поэтому их называют зародышевой плазмой, в отличие от всех других клеток, составляющих соматоплазму. Однако как зародышевая плазма, так и соматоплазма происходят из оплодотворенного яйца (зиготы), давшей начало новому организму. Следовательно, в своей основе они одинаковы. Факторы, определяющие судьбу клеток (какие клетки станут половыми, а какие — соматическими), до настоящего времени не установлены. Однако в конечном итоге половые клетки приобретают достаточно четкие отличия, возникающие в процессе гаметогенеза.
У всех позвоночных и некоторых беспозвоночных первичные половые клетки возникают вдали от гонад и мигрируют к гонадам зародыша – яичнику или семеннику — с током крови, с пластами развивающихся тканей или посредством амебоидных движений. В гонадах из них образуются зрелые половые клетки. Ко времени развития гонад сома и зародышевая плазма функционально уже обособлены, и, начиная с этого времени, на протяжении всей жизни организма половые клетки практически независимы от каких бы то ни было воздействий сомы. Поэтому признаки, приобретаемые организмом на протяжении его жизни, не оказывают влияния на его половые клетки.
Мейоз. Важное биологическое значение имеет центральное событие гаметогенеза — мейоз. Это особый способ клеточного деления, обусловливающий возникновение гаплоидных клеток и характеризующийся перекомбинированием наследственного материала между гомологичными хромосомами. Мейоз открыт В. Флемингом (1882) у животных. Мейоз начинается после репликации ДНК в предшествующей ему интерфазе. Отличительная особенность клеточного деления посредством мейоза — сложная и сильно растянутая во времени профаза.
Мейоз состоит из двух последовательных делений. Цитогенетический результат мейоза (образование гаплоидных клеток и перекомбинирование наследственного материала) определяется первым (редукционным) делением (рис. 48). Оно включает 4 фазы: профазу, метафазу, анафазу и телофазу. Профаза первого деления мейоза подразделяется на 5 стадий. На первой стадии, именуемой стадией лептотены (стадией тонких нитей), происходит спирализация ДНК ооцита, в ходе которой появляются плотные интенсивно окрашиваемые тельца. Формирующиеся хромосомы имеют вид нитей с утолщениями по длине. На последующей стадии (стадии зиготены) продолжается спирализация ДНК, а гомологичные хромосомы сближаются (рис.48) и образуют пары (синапсис хромосом). На третьей стадии (стадии пахитены) из-за продолжающейся спирализации хромосомы утолщаются до такой степени, что в ооцитах уже видны попарно расположенные гомологичные хромосомы (биваленты). Поскольку каждая гомологичная хромосома состоит из двух хроматид, то бивалент (комплекс из пары гомологичных хромосом), представленный четырьмя хроматидами, получил название тетрады. На стадии пахитены начинается кроссинговер — взаимный обмен одним или несколькими генами между спаренными (гомологичными) хромосомами в результате разрыва хроматид и воссоединения их участков в другом порядке.
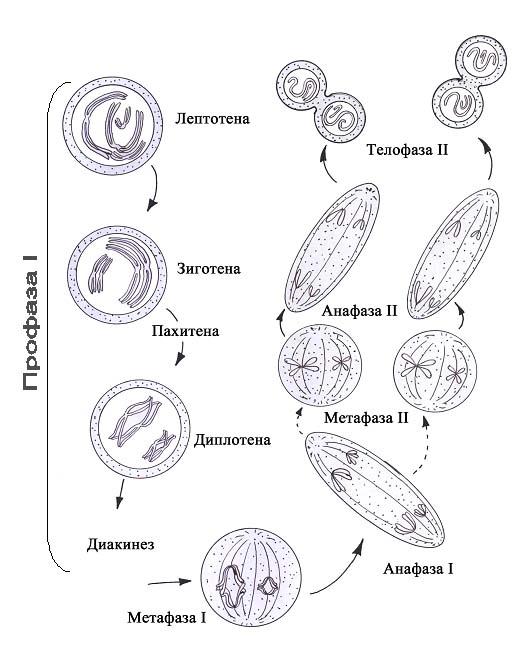
Рис. 48. Схема мейоза
Места контакта (хиазмы) сохраняются и в следующей стадии диплотены. В местах контакта мужской и женской хроматид ослабляется связь между генами и происходит обмен гомологичными генами между конъюгирующими (гомологичными) хромосомами. В стадии диплотены происходит дальнейшая спирализация хромосом, но в ооцитах многих животных, накапливающих желток, хромосомы, наоборот, деконденсируются и приобретают вид «ламповых щёток» (фаза диктиотены). Такое разрыхление хромосом сопровождается активизацией процесса синтеза РНК и белка (что необходимо для накопления ооцитом I желтка). У насекомых хромосомы типа «ламповых щёток» могут существовать год и более, у человека — 12-50 лет. На стадии диакинеза (расхождение двойных нитей) уменьшается число хиазм, парные хромосомы частично расходятся, начинает образовываться веретено деления. В метафазе I, следующей за длинной и сложной профазой I, пары хромосом (биваленты) выстраиваются по экватору клетки, образуя экваториальную пластинку. В этой фазе можно определить число хиазм (рис. 48). В среднем в ооците I женщины их насчитывается 42-50. В анафазе I к полюсам расходятся гомологичные хромосомы (диады), состоящие каждая из двух хроматид. В телофазе I происходит цитотомия и образуются один ооцит II и первое редукционное тельце либо два сперматоцита II порядка с гаплоидным набором хромосом. После этого начинается укороченная интерфаза и далее следует фактически метафаза второго (эквационного) деления мейоза. Хромосомы (диады, т.к. состоят из двух хроматид) выстраиваются по экватору клетки. В анафазе II к полюсам отходит по одной хроматиде из каждой хромосомы (рис. 48). В телофазе II образуются яйцеклетка и второе направительное тельце либо 4 сперматиды, содержащие гаплоидный набор хромосом, причём каждая хромосома состоит только из одной хроматиды. Примечательно, что второе (эквационное) деление мейоза завершается только после выхода ооцита II из яичника (после овуляции) и для его завершения необходимо, чтобы произошло оплодотворение клетки сперматозоидом (рис.45). Последние стадии оогенеза удаётся воспроизводить вне организма женщины в искусственной среде. Это позволяет осуществлять оплодотворение яйцеклетки in vitro (в пробирке). Яйцеклетку извлекают из яичника хирургическим путём ещё до овуляции и переносят в питательную среду со сперматозоидами, где после оплодотворения начинается развитие зиготы. На стадии 8-16 бластомеров зародыш переносится в матку женщины-реципиента. Поддерживать развитие зародыша в искусственной среде можно и далее, однако конечного результата при этом достичь пока не удаётся.
Мейоз, имеющий место в процессе гаметогенеза, обладает важным общебиологическим значением: случайное расхождение отцовских и материнских хромосом в процессе мейоза, а также кроссинговер обеспечивают появление новых вариантов комбинации генов, не встречавшихся у родительских форм. Этим существенно расширяется естественный материал для эволюционного процесса.
studfile.net
