Физиология — Координационная деятельность
Координационная деятельность
Координационная деятельность (КД) ЦНС представляет собой согласованную работу нейронов ЦНС, основанную на взаимодействии нейронов между собой.
Функции КД:
1) обеспечивает четкое выполнение определенных функций, рефлексов;
2) обеспечивает последовательное включение в работу различных нервных центров для обеспечения сложных форм деятельности;
3) обеспечивает согласованную работу различных нервных центров.
Основные принципы КД ЦНС и их нейронные механизмы.
1. Принцип иррадиации. При возбуждении небольших групп нейронов возбуждение распространяется на значительное количество нейронов.
2. Принцип конвергенции. При возбуждении большого количества нейронов возбуждение может сходиться к одной группе нервных клеток.
3. Принцип реципрокности – согласованная работа нервных центров, особенно у противоположных рефлексов (сгибание, разгибание и т. д.
4. Принцип доминанты. Доминанта – господствующий очаг возбуждения в ЦНС в данный момент. Доминанта лежит в основе формирования условного рефлекса.
5. Принцип обратной связи. Различают два вида обратной связи:
1) положительная обратная связь, вызывающая усиление ответной реакции со стороны нервной системы.
Лежит в основе порочного круга, который приводит к развитию заболеваний;
2) отрицательная обратная связь, снижающая активность нейронов ЦНС и ответную реакцию. Лежит в основе саморегуляции.
6. Принцип субординации. В ЦНС существует определенная подчиненность отделов друг другу, высшим отделом является кора головного мозга.
7. Принцип взаимодействия процессов возбуждения и торможения. ЦНС координирует процессы возбуждения и торможения: оба процесса способны к конвергенции, процесс возбуждения и в меньшей степени торможения способны к иррадиации. Торможение и возбуждение связаны индукционными взаимоотношениями. Процесс возбуждения индуцирует торможение, и наоборот.
1) последовательная. Процесс возбуждения и торможения сменяют друг друга по времени;
2) взаимная. Одновременно существует два процесса – возбуждения и торможения.
Координационная деятельность ЦНС обеспечивает четкое взаимодействие между отдельными нервными клетками и отдельными группами нервных клеток.
36. Основные принципы координационной деятельности цнс: реципрокности,
облегчения, окклюзии, обратной связи, общего «конечного» пути, доминанты.
Рефлекторная реакция в большинстве случаев осуществляется не одной, а целой группой рефлекторных дуг и нервных центров. Координация рефлекторной деятельности это такое взаимодействие нервных центров и проходящих по ним нервных импульсов, которое обеспечивает согласованную деятельность органов и систем организма. Она осуществляется с помощью следующих процессов:
1. Временное и пространственное облегчение.
Это усиление рефлекторной реакции при
действии ряда последовательных
раздражителей или одновременном их
воздействии на несколько рецептивных
полей. Объясняется явлением суммации
в нервных центрах.
Временное и пространственное облегчение.
Это усиление рефлекторной реакции при
действии ряда последовательных
раздражителей или одновременном их
воздействии на несколько рецептивных
полей. Объясняется явлением суммации
в нервных центрах.
2. Окклюзия явление противоположное облегчению. Когда рефлекторная реакция на два или более сверхпороговых раздражителя меньше, чем ответы на их раздельное воздействие. Оно связано с конвергенцией нескольких возбуждающих импульсов на одном нейроне.
3.
Принцип общего конечного пути. Разработан
Ч. Шеррингтоном. В основе его лежит
явление конвергенции. Согласно этому
принципу на одном эфферентном мотонейроне
могут образовывать синапсы нескольких
афферентных, входящих в несколько
рефлекторных дуг. Этот нейрон называется
общим конечным путем и участвует в
нескольких рефлекторных реакциях. Если
взаимодействие этих рефлексов приводит
к усилению общей рефлекторной реакции,
такие рефлексы называются союзными.
Если же между афферентными сигналами
происходит борьба за мотонейрон —
конечный путь, то антагонистическими.
4. Реципрокное торможение. Обнаружено Ч. Шеррингтоном. Это явление
торможения одного центра в результате возбуждения другого. Т.е. в этом случае тормозится антагонистический центр. Например при возбуждении центров сгибания левой ноги, по реципрокному механизму тормозятся центры мышц разгибателей этой же ноги и центры сгибателей правой. В реципрокных взаимоотношениях находятся центры вдоха и выдоха продолговатого мозга, центры сна и бодрствования и т.д.
5.
Принцип доминанты. Открыт А.А. Ухтомским.
Доминанта — это преобладающий очаг
возбуждения в ЦНС, подчиняющий себе
другие НЦ. Доминантный центр обеспечивает
комплекс рефлексов, которые необходимы
в данный момент для достижения определенной
цели. При некоторых условиях возникают
питьевая, пищевая, оборонительная,
половая и др. доминанты. Свойствами
доминантного очага являются повышенная
возбудимость, стойкость возбуждения,
высокая способность к суммации,
инертность.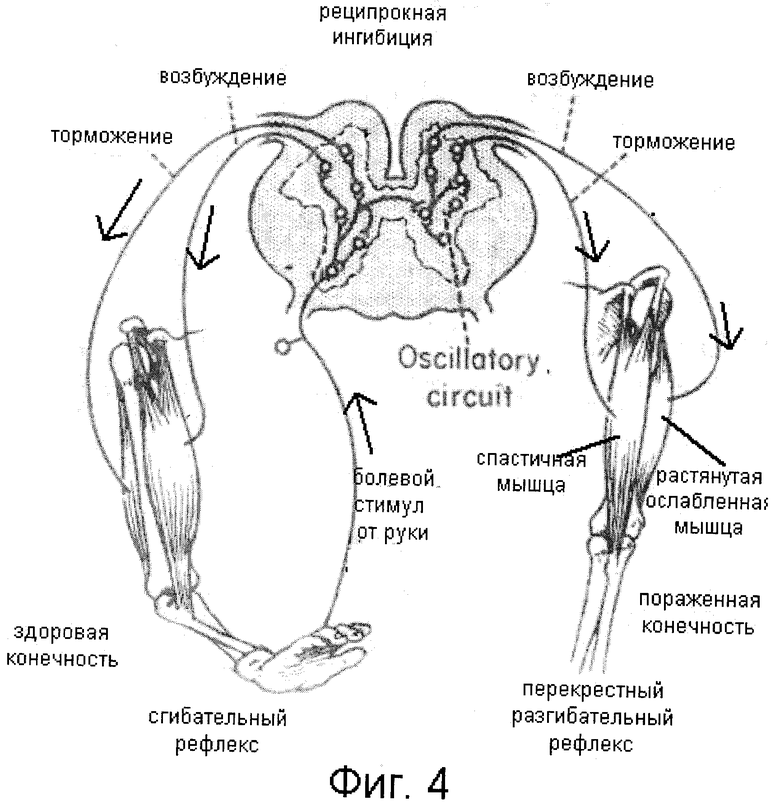 Эти свойства обусловлены
явлениями облегчения, иррадиации, с
одновременным повышением активности
вставочных тормозных нейронов, которые
тормозят нейроны других центров.
Эти свойства обусловлены
явлениями облегчения, иррадиации, с
одновременным повышением активности
вставочных тормозных нейронов, которые
тормозят нейроны других центров.
6. Принцип обратной афферентации. Результаты рефлекторного акта воспринимаются нейронами обратной афферентации и информация от них поступает обратно в нервный центр. Там они сравниваются с параметрами возбуждения и рефлекторная реакция корректируется.
Основные принципы координационной деятельности цнс
В основе
координационной деятельности ЦНС лежит
взаимодействие между процессами
возбуждения и торможения. О существовании
возбуждения в нервах, мышцах, в ЦНС было
известно давно. Торможение в ЦНС было
открыто И.М.Сеченовым (1862 г.) в опытах на
лягушках и получило название «Сеченовское
торможение». Он определял время
сгибательного рефлекса (по Тюрку),
погружая лапку лягушки в кислоту, а
затем на зрительные бугры накладывал
кристаллик поваренной соли. После
наложения кристаллика происходило
удлинение времени рефлекса или рефлекс
полностью затормаживался, а после снятия
кристаллика соли и промывания этого
участка мозга водой время рефлекса
восстанавливалось до исходного уровня.
Согласованная (координационная)
деятельность обеспечивается за счет
ряда механизмов:
После
наложения кристаллика происходило
удлинение времени рефлекса или рефлекс
полностью затормаживался, а после снятия
кристаллика соли и промывания этого
участка мозга водой время рефлекса
восстанавливалось до исходного уровня.
Согласованная (координационная)
деятельность обеспечивается за счет
ряда механизмов:
1) Принцип доминанты. Он был сформулирован А.А.Ухтомским как основной принцип работы нервных центров. Доминантный (или господствующий) очаг возбуждения характеризуется следующими свойствами: повышенной возбудимостью; инертностью (стойкостью) возбуждения, т.е. может сохраняться длительное время; способностью к суммации возбуждений, притягивая на себя возбуждение с других центров; способностью тормозить субдоминантные очаги возбуждения других нервных центров.
2) Принцип окклюзии.
Этот принцип противоположен
пространственному облегчению или
суммации, и он заключается в том, что
два афферентных входа совместно
возбуждают меньшую группу мотонейронов
по сравнению с эффектами при раздельной
их активации. Причина окклюзии состоит
в том, что афферентные входы в силу
конвергенции отчасти адресуются к одним
и тем же мотонейронам, которые
затормаживаются при активации обоих
входов одновременно. Явление окклюзии
проявляется в случаях применения сильных
афферентных раздражений.
Причина окклюзии состоит
в том, что афферентные входы в силу
конвергенции отчасти адресуются к одним
и тем же мотонейронам, которые
затормаживаются при активации обоих
входов одновременно. Явление окклюзии
проявляется в случаях применения сильных
афферентных раздражений.
3) Принцип обратной связи. Процессы саморегуляции в организме в полном объеме могут осуществляться только в том случае, когда функционирует канал обратной связи. За счет импульсов, поступающих по этому каналу, происходит оценка правильности исполнения поставленной задачи, а если она не выполнена, то вносятся коррекции для достижения результата.
Велико значение
механизмов обратной связи в поддержании
гомеостаза. Так, например, поддержание
постоянного уровня кровяного давления
осуществляется за счет изменения
импульсной активности барорецепторов
сосудистых рефлексогенных зон, в
результате чего измененяеся тонус
вазомоторных симпатических нервов и
таким образом нормализуют кровяное
давление.
4) Принцип
реципрокности (сочетанности, сопряженности,
взаимообусловленности). Он отражает
характер отношений между центрами
ответственными за осуществление
противоположных функций (вдоха и
глотания, выдоха и выдоха, сгибания и
разгибания конечностей и т.д.). Например,
активация проприорецепторов мышцы-сгибателя
одновременно возбуждает центр мышц
сгибателей и тормозит центр мышц
разгибателей. Реципрокное торможение
играет важную роль в координации
двигательных актов. Реципркные отношения
имеют динамический характер (о чем
говорил еще Введенский), а Шеррингтон
эти отношения рассматривал как статические
явления. Опытами П.К.Анохина с перекрестным
подшиванием сухожилий сгибателей к
разгибателям и наоборот было установлено,
что через 6–8 месяцев мышцы сгибатели
начинают выполнять функцию разгибателей,
а разгибатели функцию сгибателей. Такая
перестройка реципрокных взаимоотношений
была бы невозможна, если реципрокные
отношения имели бы строго раз и навсегда
зафиксированный (статический) характер.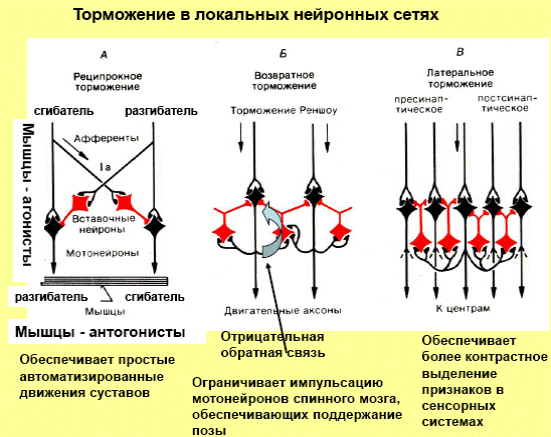
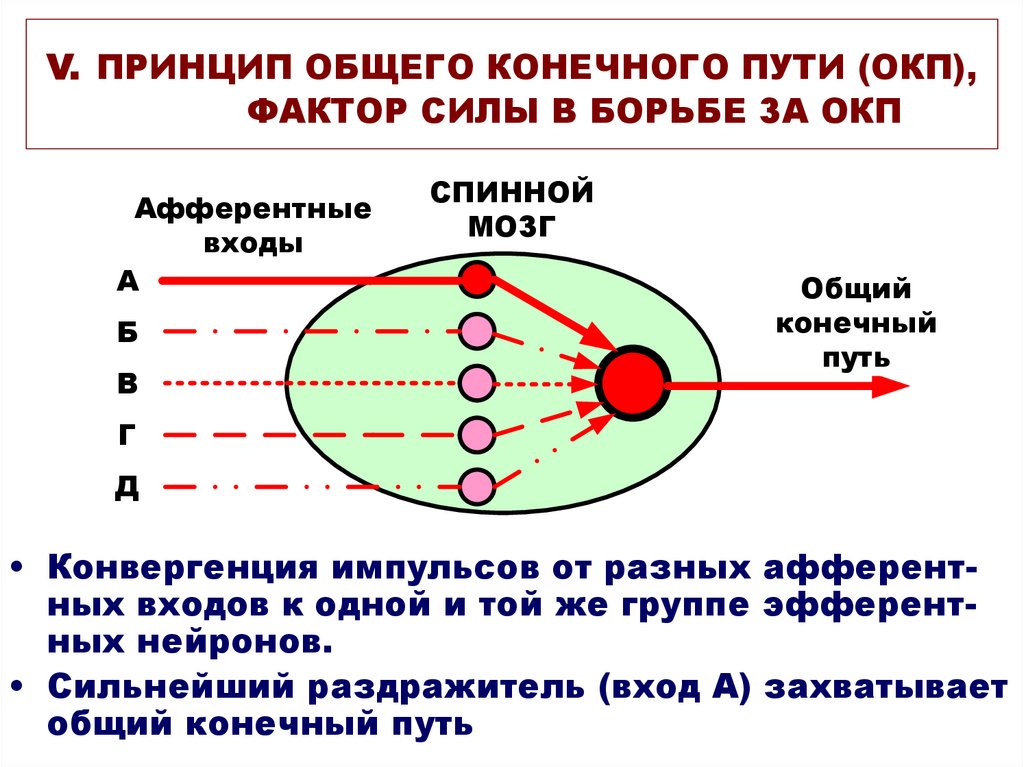
5) Принцип общего конечного пути. Эффекторные нейроны ЦНС, например, мотонейроны спинного мозга, могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого числа афферентных и промежуточных нейронов, для которых они являются конечным путем (путем от ЦНС к эффектору). Например, на мотонейронах передних рогов спинного мозга, иннервирующих мускулатуру конечности, оканчиваются волокна афферентных нейронов, нейронов пирамидного тракта и экстрапирамидной системы (ядер мозжечка, ретикулярной формации и многих других структур)
6) Явления конвергенции
—
схождение
нервных импульсов на одни и те же
центральные нейроны. Такая особенность
зависит не только от функциональных
свойств центров, но обусловлено также
количественными соотношениями
периферических рецепторных и промежуточных
центральных нейронов. Это соотношение
составляет примерно 10:1. Явления
конвергенции играют решающее значение
при формировании общего конечного пути.
Такая особенность
зависит не только от функциональных
свойств центров, но обусловлено также
количественными соотношениями
периферических рецепторных и промежуточных
центральных нейронов. Это соотношение
составляет примерно 10:1. Явления
конвергенции играют решающее значение
при формировании общего конечного пути.
7) Явления дивергенции — процесс противоположный конвергенции, т.е. импульсы, поступающие в ЦНС, распространяются (иррадиируют) на соседние участки.
8) Субординационные взаимоотношения — соподчинение, т.е. вышележащие отделы ЦНС оказывают свое регулирующее влияние на нижележащие отделы.
Координация функций в ЦНС | Кинезиолог
Принципы координации в ЦНС
Координация – это согласование и сопряжение нервных процессов, характерное для деятельности центральной нервной системы (ЦНС).
1. Принцип реципрокной (сопряженной, взаимоисключающей) иннервации.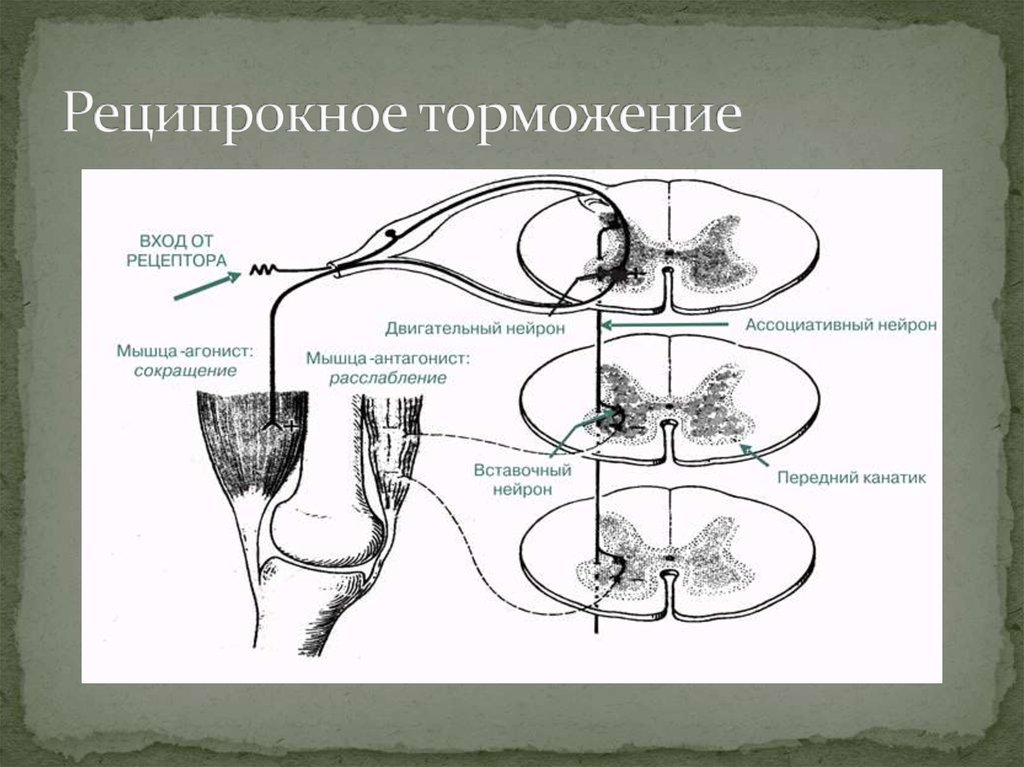
2. Принцип общего конечного пути (принцип конвергенции, «воронка Ч. Шеррингтона»).
3. Принцип доминанты.
4. Принцип временнОй связи.
5. Принцип саморегуляции (прямая и обратная связи).
6. Принцип иерархии (соподчинение).
1. Принцип реципрокной (сопряженной, взаимоисключающей) иннервации
Принцип реципропной иннервации мышц-антагонистов впервые был обнаружен в 1896 году выдающимся отечественным физиологом Н.Е. Введенским, учеником И.М. Сеченова.
Сокращение сгибателя вызывает понижение тонуса разгибателя на той же стороне, а с противоположной
стороны — наоборот: может вызывать повышение тонуса разгибателя.
На реципрокном принципе основан рефлекс шагания. Таким образом, ходьба — это условнорефлекторная, основанная на принципе реципрокной иннервации, циклическая двигательная деятельность ног.
Вывод:
Возбуждение сгибателя вызывает сопряженное торможение и расслабление разгибателя: происходит перекрестный разгибательный рефлекс.
2. Принцип общего конечного пути (принцип конвергенции)
Этот принцип был открыт и исследован выдающимся английским физиологом сэром Ч.С. Шеррингтоном (Charles Scott Sherrington) в 1896 году.
Он установил, что в нервных центрах количество афферентных (приносящих) клеток намного больше, чем количество эфферентных (выносящих) нейронов, несущих возбуждение к мышцам. Получается, что между нейронами идет борьба «за общий конечный путь», т.е. за то, чтобы передать своё возбуждение на эффентные нейроны. Этот принцип получил также образное наименование «воронки Шеррингтона».
3. Принцип доминанты
Доминанта (от лат. «господствовать») — это временно господствующий рефлекс, подчиняющий себе дуги прочих рефлексов. Доминанта существует в виде устойчивого очага возбуждения, подчиняющего себе другие возбуждённые очаги.
Доминанта может быть гуморальной, или её можно вызвать искусственно, если вызвать деполяризацию участка головного мозга с помощью химического или электрического воздействия.
Примеры доминанты:
Попытки лягушки снять себя с крючка.
Особенности доминантного очага (центра) :
— повышенная возбудимость,
— повышенная стойкость (устойчивость к тормозным воздействиям),
— тормозное воздействие на другие возбуждающиеся очаги,
— способность к суммации возбуждения с соседних участков,
— длительность существования данного возбуждённого очага,
— инерция, т.е. длительное удержание возбуждённого состояния после прекращения первоначального возбуждения и сопротивление тормозному воздействию.
Доминанта была открыта в 1924 году А.А. Ухтомским, крупным отечественным физиологом, учеником другого крупного физиологи — Н.Е. Введенского.
Суть этого явления заключается в том, что если существует доминантный очаг, имеющий возбуждение, то любое другое возбуждение будет усиливать реакцию именно этого доминантного очага. И рефлекторный ответ будет соответствовать именно доминантному очагу (доминантному нервному центру), а не раздражителю. Можно сказать, что доминанта нарушает протекание классических условных и безусловных рефлексов. Кроме того, доминантный очаг тормозит все другие центры и подавляет их возбуждение. Таким образом, доминанта как бы фильтрует возбуждение, приходящее из разных источников, т.к. тормозит все посторонние ненужные импульсы.
Можно сказать, что доминанта нарушает протекание классических условных и безусловных рефлексов. Кроме того, доминантный очаг тормозит все другие центры и подавляет их возбуждение. Таким образом, доминанта как бы фильтрует возбуждение, приходящее из разных источников, т.к. тормозит все посторонние ненужные импульсы.
В 1960-е годы В.С. Русинов получил искусственную доминанту путем слабого электрического раздражения 6-го слоя коры больших полушарий головного мозга.
Иногда в основе доминанты лежит снижение лабильности (подвижности нервных процессов).
Формы доминанты
1. Чувствительная (сенсорная).
2. Двигательная.
По механизму:
1. Рефлекторная.
2. Гуморальная (голодовая, половая).
По уровню расположения:
1. Спинальная (спинной мозг).
2. Бульбарная (продолговатый мозг).
3. Мезэнцефальная (средний мозг).
4. Диэнцефальная (промежуточный мозг).
5. Кортикальная (корковая).
4. Принцип временной связи
Принцип временной связи
Высшая форма временной связи — условный рефлекс.
5. Принцип саморегуляции (прямая и обратная связи)
Прямые и обратные связи — это пути влияния управляющего объекта на управляемый объект. Соответственно, влияние может быть прямым и обратным.
Обратные связи, в свою очередь, делятся на положительные (усиливающие) и отрицательные (ослабляющие).
6. Принцип иерархии (соподчинение)
Принцип иерархии очень прост — нижележащие структуры подчиняются вышележащим. Это означает, что вышележащие структуры умеют как подгонять, так и тормозить нижележащие структуры.
Существует также функциональная иерархия. Так, высшее место в иерархии безусловных рефлексов занимает оборонительный рефлекс, затем — пищевой рефлекс, потом половой. Но в ряще случаев лидерство может захватывать половой рефлекс, оттесняя на второй план пищевое поведение и даже инстинкт самосохранения.
Дополнительные материалы
Принципы координационной деятельности мозга
Принципы координации в деятельности ЦНС.
 — Студопедия
— СтудопедияВ условиях физиологической нормы работа всех органов и систем тела является согласованной: на воздействия из внешней и внутренней среды организм реагирует как единое целое. Согласованное проявление отдельных рефлексов, обеспечивающих выполнение целостных рабочих актов, носит название координации.
Явления координации играют важную роль в деятельности двигательного аппарата. Координация таких двигательных актов, как ходьба или бег, обеспечивается взаимосвязанной работой нервных центров.
За счет координированной работы нервных центров осуществляется совершенное приспособление организма к условиям существования. Это происходит не только за счет деятельности двигательного аппарата, но и за счет изменений вегетативных функций организма (процессов дыхания, кровообращения, пищеварения, обмена веществ и т.д.).
Установлен ряд общих закономерностей — принципов координации:
— принцип конвергенции;
— принцип иррадиации возбуждения;
— принцип реципрокности;
— принцип последовательной смены возбуждения торможением и торможения возбуждением;
— феномен «отдачи»;
— цепные и ритмические рефлексы;
— принцип общего конечного пути;
— принцип обратной связи;
— принцип доминанты.
Разберем некоторые из них.
Принцип конвергенции. Этот принцип установлен английским физиологом Шеррингтоном. Импульсы, при ходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным и эфферентным нейронам. Конвергенция нервных импульсов объясняется тем, что афферентных нейронов в несколько раз больше, чем эфферентных, поэтому афферентные нейроны образуют на телах и дендритах эфферентных и вставочных нейронов многочисленные синапсы.
Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры.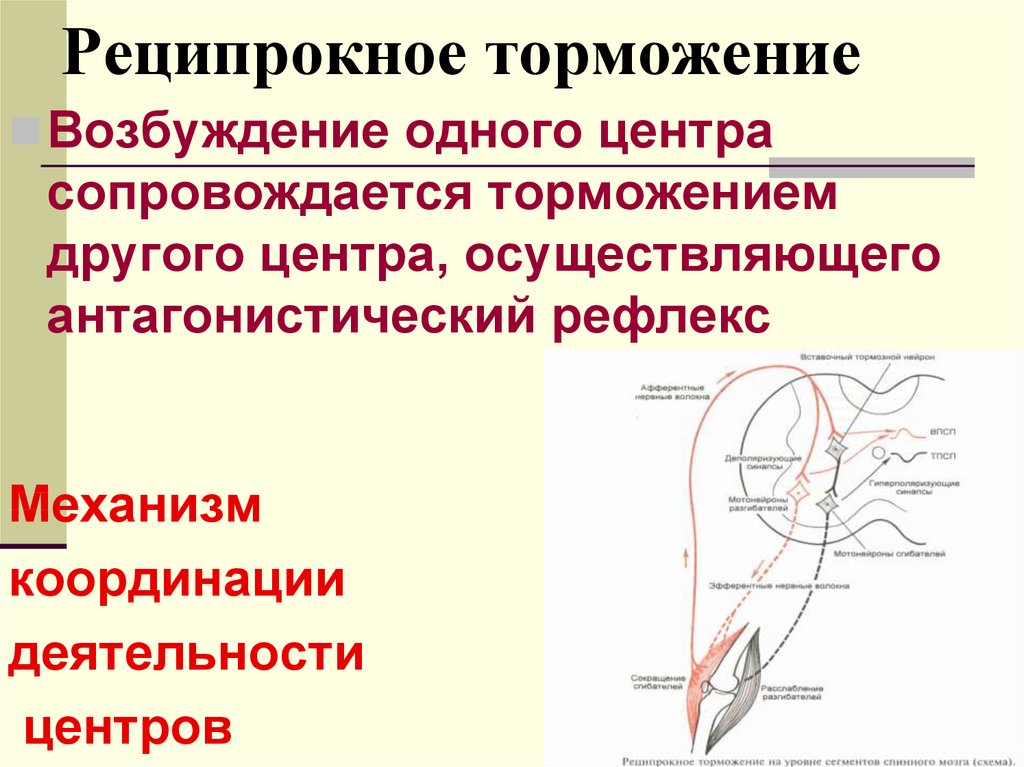
Принцип реципрокности (сопряженности) в работе нервных центров. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов — сгибателей и разгибателей конечностей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы.
Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное), но может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д.
Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр.
Принцип общего конечного пути. Этот принцип связан с особенностью строения центральной нервной системы. Эта особенность, как уже указывалось, состоит в том, что афферентных нейронов в несколько раз больше, чем эфферентных, в результате чего различные афферентные импульсы сходятся к общим выходящим путям.
Количественные соотношения между нейронами схематически можно представить в виде воронки: возбуждение вливается в центральную нервную систему через широкий раструб (афферентные нейроны) и вытекает из нее через узкую трубку (эфферентные нейроны). Общими путями могут быть не только конечные эфферентные нейроны, но и вставочные.
Импульсы, сходящиеся в общем пути, «конкурируют» друг с другом за использование этого пути. Так достигается упорядочение рефлекторного ответа, соподчинение рефлексов и затормаживание менее существенных. Вместе с тем организм получает возможность реагировать на различные раздражения из внешней и внутренней среды при помощи сравнительно небольшого количества исполнительных органов.
Принцип обратной связи. Этот принцип изучен И. М. Сеченовым, Шеррингтоном, П. К. Анохиным и рядом других исследователей. При рефлекторном сокращении скелетных мышц возбуждаются проприорецепторы. От проприорецепторов нервные импульсы несущие информацию о характеристиках этого мышечного сокращения вновь поступают в центральную нервную систему. Этим контролируется точность совершаемых движений. Подобные афферентные импульсы, возникающие в организме в результате рефлекторной деятельности органов и тканей (эффекторов), получили название вторичных афферентных импульсов, или обратной связи.
Обратные связи могут быть положительными и отрицательными. Положительные обратные связи способствуют усилению рефлекторных реакций, отрицательные — их угнетению. За счет положительных и отрицательных обратных связей осуществляется, например, регуляция относительного постоянства величины артериального давления.
Принцип доминанты. Принцип доминанты сформулирован А. Л. Ухтомским, Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта — это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Л. Ухтомским, Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта — это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Доминантный очаг возбуждения характеризуется следующими основными свойствами:
— повышенной возбудимостью;
— стойкостью возбуждения;
— способностью к суммированию возбуждения;
— инерцией, доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения.
Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы из других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется.
Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека.
Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения.
Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, а также эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
Рефлекторная реакция осуществляется целой группой рефлекторных дуг и нервных центров. Временное и пространственное облегчение. Это усиление рефлекторной реакции при действии ряда последовательных раздражителей или одновременном их воздействии на несколько рецептивных полей. Объясняется явлением суммации в нервных центрах. Окклюзияявление противоположное облегчению. Когда рефлекторная реакция на два или более сверхпороговых раздражителя меньше, чем ответы на их раздельное воздействие. Оно связано с конвергенцией нескольких возбуждающих импульсов на одном нейроне. Принцип общего конечного пути. Разработан Ч. Шеррингтоном. В основе его лежит явление конвергенции. Согласно этому принципу на одном эфферентном мотонейроне могут образовывать синапсы нескольких афферентных, входящих в несколько рефлекторных дуг. Этот нейрон называется общим конечным путем и участвует в нескольких рефлекторных реакциях. Реципрокное торможение. Обнаружено Ч. Шеррингтоном. Это явление торможения одного центра в результате возбуждения другого. Т.е. в этом случае тормозится антагонистический центр. В реципрокных взаимоотношениях находятся центры вдоха и выдоха продолговатого мозга, центры сна и бодрствования и т.д. Принцип доминанты. Открыт А.А. Ухтомским. Доминанта — это преобладающий очаг возбуждения в ЦНС, подчиняющий себе другие НЦ. Доминантный центр обеспечивает комплекс рефлексов, которые необходимы в данный момент для достижения определенной цели. При некоторых условиях возникают питьевая, пищевая, оборонительная, половая и др. доминанты. Свойствами доминантного очага являются повышенная возбудимость, стойкость возбуждения, высокая способность к суммации, инертность. Принцип обратной афферентации. Результаты рефлекторного акта воспринимаются нейронами обратной афферентации и информация от них поступает обратно в нервный центр. Там они сравниваются с параметрами возбуждения и рефлекторная реакция корректируется.
24. Роль спинного мозга в процессах регуляции соматических и вегетативных функций организма. Клинически важные спинальные рефлексы. Проводниковая функция спинного мозга.Спинной мозг находится в позвоночном канале и выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями. Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, т.е. афферентными, передние двигательными, эфферентными. Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все его нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов вегетативной нервной системы. Все рефлексы спинного мозга делятся на соматические, т.е. двигательные и вегетативные. Соматические рефлексы делятся на сухожильные и кожные. Сухожильные рефлексы возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и α-мотонейронов спинного мозга. В результате возникает сокращение мышц, в первую очередь разгибателей.
25. Продолговатый мозг и мост, участие их центров в процессах саморегуляции функций.Основными функциями продолговатого мозга являются проводниковая, рефлекторная и ассоциативная. Первая осуществляется проводящими путями, проходящими через него. Вторая, нервными центрами. В ромбовидной ямке продолговатого мозга находятся ядра 10, 11, 12 пар черепно-мозговых нервов, а также ретикулярная формация. Рефлекторные функции делятся на соматические и вегетативные. Соматическими являются статические рефлексы продолговатого мозга, относящиеся к позно-тоническим или рефлексам позы. Мост имеет тесные функциональные связи со средним мозгом. Эти отделы ствола мозга также осуществляют проводниковую и рефлекторную функции. Проводниковая обеспечивается восходящими и нисходящими путями, идущими через них. Рефлекторная их нервными центрами. Нейроны моста формируют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательную часть ядер тройничного нерва и его чувствительное ядро. Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте: |
 Координация рефлекторной деятельности обеспечивает согласованную деятельность органов и систем организма. Она осуществляется с помощью следующих процессов:
Координация рефлекторной деятельности обеспечивает согласованную деятельность органов и систем организма. Она осуществляется с помощью следующих процессов: Если взаимодействие этих рефлексов приводит к усилению общей рефлекторной реакции, такие рефлексы называются союзными. Если же между афферентными сигналами происходит борьба за мотонейрон — конечный путь, то антагонистическими.
Если взаимодействие этих рефлексов приводит к усилению общей рефлекторной реакции, такие рефлексы называются союзными. Если же между афферентными сигналами происходит борьба за мотонейрон — конечный путь, то антагонистическими.
 Мотонейроны, в зависимости от функционального значения, разделяются на альфа- и гамма-мотонейроны. К α-мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных, т.е. рецепторных мышечных клеток. Тела α-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает. Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС.
Мотонейроны, в зависимости от функционального значения, разделяются на альфа- и гамма-мотонейроны. К α-мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных, т.е. рецепторных мышечных клеток. Тела α-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает. Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной (объяснение). Спинальные нервные центры находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится. Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.Проводниковая функция состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие.
Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной (объяснение). Спинальные нервные центры находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится. Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.Проводниковая функция состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие. Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища. К восходящим путям относятся:Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности. Нисходящие пути также образованы несколькими трактами. ортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры к α-мотонейронам спинного мозга.
Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища. К восходящим путям относятся:Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности. Нисходящие пути также образованы несколькими трактами. ортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры к α-мотонейронам спинного мозга.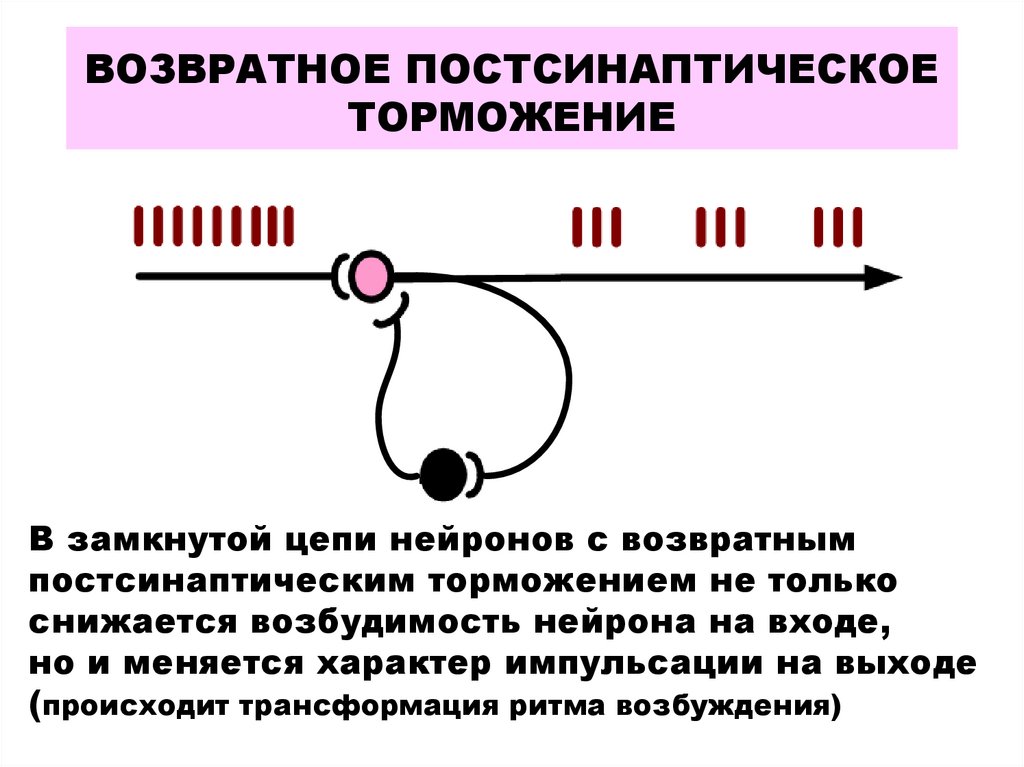 За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга к α-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к α-мотонейронам мышц разгибателей. За счет этих двух путей регулируется тонус соответствующих мышц при изменениях положения тела.
За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга к α-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к α-мотонейронам мышц разгибателей. За счет этих двух путей регулируется тонус соответствующих мышц при изменениях положения тела.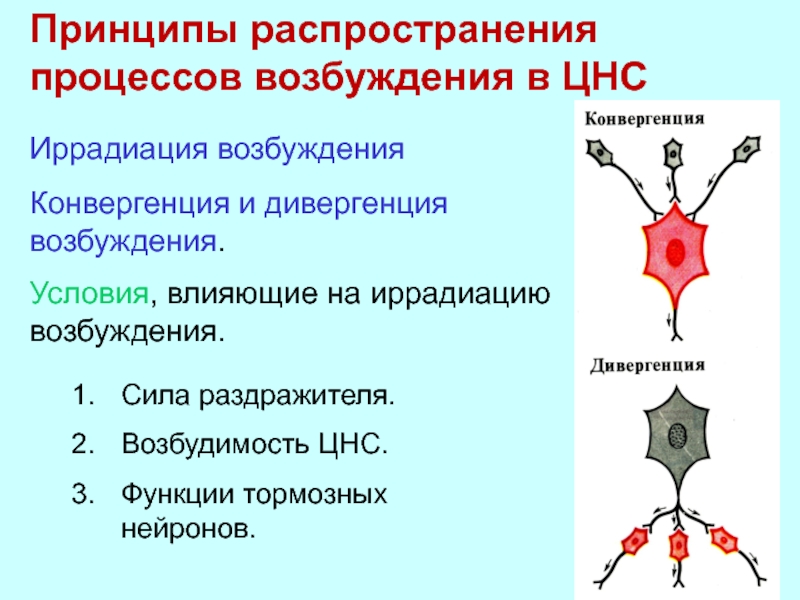 Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер. От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальныне тракты. Рефлексы возникают тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышается тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция. При повороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности. В продолговатом мозге находятся жизненно важные центры. К ним относятся дыхательный, сосудодвигательный центры и центр регуляции сердечной деятельности. Первый обеспечивает смену фаз дыхания, второй — тонус периферических сосудов, третий регуляцию частоты и силы сердечных сокращений. В области ядер блуждающего нерва находятся центры слюноотделения, секреции желудочных, кишечных желез, поджелудочной железы и печени.
Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер. От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальныне тракты. Рефлексы возникают тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышается тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция. При повороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности. В продолговатом мозге находятся жизненно важные центры. К ним относятся дыхательный, сосудодвигательный центры и центр регуляции сердечной деятельности. Первый обеспечивает смену фаз дыхания, второй — тонус периферических сосудов, третий регуляцию частоты и силы сердечных сокращений. В области ядер блуждающего нерва находятся центры слюноотделения, секреции желудочных, кишечных желез, поджелудочной железы и печени. Здесь же расположены центры регуляции моторики пищеварительного канала. Важной функцией продолговатого мозга является формирование защитных рефлексов. В нем находятся рвотный центр, центры кашля, чихания, смыкания век и слезотечения при раздражении роговицы. Здесь расположены бульбарные отделы центров, участвующих в организации пищевых рефлексов — сосания, жевания, глотания. В продолговатом мозге происходит первичный анализ ряда сенсорных сигналов. В частности, в нем расположены ядра слухового нерва, верхнее вестибулярное ядро, а к ядрам языкоглоточного нерва поступают сигналы от вкусовых рецепторов. От рецепторов кожи лица они идут к ядрам тройничного нерва.
Здесь же расположены центры регуляции моторики пищеварительного канала. Важной функцией продолговатого мозга является формирование защитных рефлексов. В нем находятся рвотный центр, центры кашля, чихания, смыкания век и слезотечения при раздражении роговицы. Здесь расположены бульбарные отделы центров, участвующих в организации пищевых рефлексов — сосания, жевания, глотания. В продолговатом мозге происходит первичный анализ ряда сенсорных сигналов. В частности, в нем расположены ядра слухового нерва, верхнее вестибулярное ядро, а к ядрам языкоглоточного нерва поступают сигналы от вкусовых рецепторов. От рецепторов кожи лица они идут к ядрам тройничного нерва.
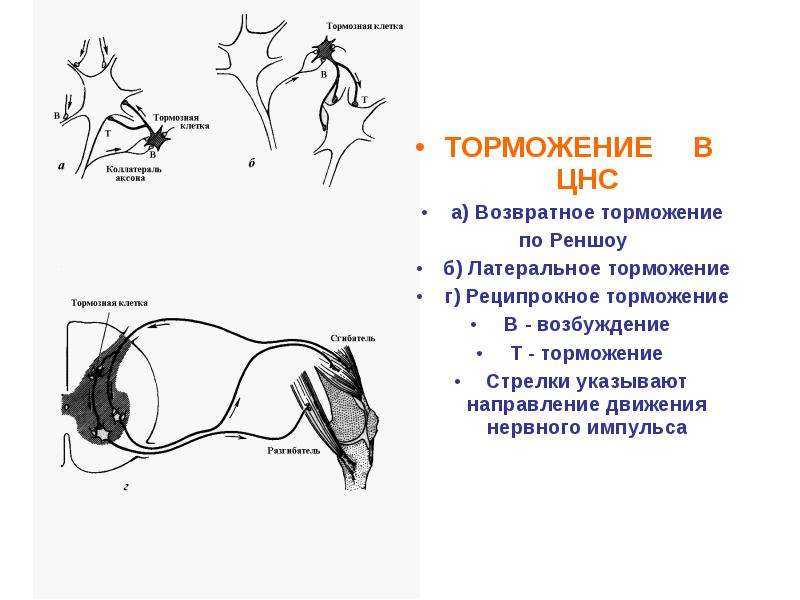
Торможение в нервных клетках и его механизмы. Принцип сопряженного торможения или реципрокности. Принцип субординации нервных центров
Торможение — особый нервный процесс, который обусловливается возбуждением и внешне проявляется угнетением другого возбуждения. Оно способно активно распространяться нервной клеткой и ее отростками. Основал учение о центральноv торможение И. М. Сеченов (1863), который заметил, что изгибающий рефлекс лягушки тормозится при химическом раздражении среднего мозга. Торможение играет важную роль в деятельности ЦНС, а именно: в координации рефлексов; в поведении человека и животных; в регуляции деятельности внутренних органов и систем; в осуществлении защитной функции нервных клеток.
Типы торможения в ЦНС
Центральное торможение распределяется по локализации на пре-и постсинаптическое;
по характеру поляризации (зарядом мембраны) — на гипер-и деполяризации;
по строению тормозных нейронных цепей — на реципрокное, или соединенное, обратное и латеральное.
Пресинаптическое торможение , как свидетельствует название, локализуется в пресинаптических элементах и связано с угнетением проведения нервных импульсов в аксональных (пресинаптических) окончаниях. Гистологическим субстратом такого торможения является аксональные синапсы. К возбуждающему аксону подходит вставной тормозной аксон, который выделяет тормозной медиатор ГАМК . Этот медиатор действует на постсинаптическую мембрану, которая является мембраной возбуждающего аксона, и вызывает в ней деполяризацию. Возникшая деполяризация тормозит вход Са2 + из синаптической щели в заключение возбуждающего аксона и таким образом приводит к снижению выброса возбуждающего медиатора в синаптическую щель, торможение реакции. Пресинаптическое торможение достигает максимума через 15-20 мс и длится около 150 мс, то есть гораздо дольше, чем постсинаптическое торможение. Пресинаптическое торможение блокируется судорожными ядами — бикулином и пикротоксин, которые являются конкурентными антагонистами ГАМК .
Постсинаптическое торможение (ГПСП) обусловлено выделением пресинаптическим окончанием аксона тормозного медиатора, который снижает или тормозит возбудимость мембран сомы и дендритов нервной клетки, с которой он контактирует. Оно связано с существованием тормозных нейронов, аксоны которых образуют на соме и дендритах клеток нервных окончаний, выделяя тормозные медиаторы — ГАМК и глицин. Под влиянием этих медиаторов возникает торможение возбуждающих нейронов. Примерами тормозных нейронов являются клетки Реншоу
в спинном мозге, нейроны грушевидные (клетки Пуркинье мозжечка), звездчатые клетки коры большого, мозга и др..
Исследованием П. Г. Костюка (1977) доказано, что постсинаптического торможения связано с первичной гиперполяризацией мембраны сомы нейрона, в основе которой лежит повышение проницаемости постсинаптической мембраны для К +. Вследствие гиперполяризации уровень мембранного потенциала удаляется от критического (порогового) уровня. То есть происходит его увеличение — гиперполяризация. Это приводит к торможению нейрона. Такой вид торможения называется гиперполяризационным.
Амплитуда и полярность ГПСП зависят от исходного уровня мембранного потенциала самого нейрона. Механизм этого явления связан с Сl + . С началом развития ТПСП Сl — входит в клетку. Когда в клетке становится его больше, чем снаружи, глицин конформирует мембрану и через открытые ее отверстия Сl + выходит из клетки. В ней уменьшается количество отрицательных зарядов, развивается деполяризация. Такой вид торможения называется деполяризационным.
Постсинаптическое торможение локальное. Развивается оно градуально, способное к суммации, не оставляет после себя рефрактерности
. Является более оперативным, четко адресованным и универсальным тормозным механизмом. По своей сути это «центральное торможение», которое было описано в свое время Ch. S. Sherrington (1906).
В зависимости от структуры тормозного нейронного цепочки, различают следующие формы постсинаптического торможения: реципрокное, обратное и латеральное, которое является собственно разновидностью обратного.
Реципрокное (сочетанное) торможение характеризуется тем, что в том случае, когда при активизации афферентов возбуждаются, например, мотонейроны мышц-сгибателей, то одновременно (на этой стороне) тормозятся мотонейроны мышц-разгибателей, действующие на этот же сустав. Происходит это потому, что афференты от мышечных веретен образуют возбуждающие синапсы на мотонейронах мышц-агонистов, а через посредство вставного тормозного нейрона — тормозные синапсы на мотонейронах мышц-антагонистов. С физиологической точки зрения такое торможение очень выгодно, поскольку облегчает движение сустава «автоматически», без дополнительного произвольного или непроизвольного контроля.
Обратное торможение. В этом случае от аксонов мотонейрона отходит одна или несколько коллатералей, которые направляются в вставных тормозных нейронов, например, клеток Реншоу. В свою очередь, клетки Реншоу образуют тормозные синапсы на мотонейроны. В случае возбуждения мотонейрона активизируются и клетки Реншоу, вследствие чего происходит гиперполяризация мембраны мотонейрона и тормозится его деятельность. Чем больше возбуждается мотонейрон, тем больше ощутимые тормозные влияния через клетки Реншоу. Таким образом, обратное постсинаптическое торможение функционирует по принципу отрицательной обратной связи. Есть предположение, что этот вид торможения требуется для саморегуляции возбуждения нейронов, а также для предотвращения их перевозбуждению и судорожным реакциям.
Латеральное торможение. Тормозная цепь нейронов характеризуется тем, что вставные тормозные нейроны влияют не только на воспаленную клетку, но и на соседние нейроны, в которых возбуждение является слабым или вовсе отсутствует. Такое торможение называется латеральным, поскольку участок торможения, который образуется, содержится сбоку (латерально) от возбужденного нейрона. Оно играет особенно важную роль в сенсорных системах, создавая явление контраста.
Постсинаптическое торможения преимущественно легко снимается при введении стрихнина, который конкурирует с тормозным медиатором (глицином) на постсинаптической мембране. Столбнячный токсин также подавляет постсинаптическое торможение, нарушая высвобождение медиатора из тормозных пресинаптических окончаний. Поэтому введение стрихнина или столбнячного токсина сопровождается судорогами, которые возникают вследствии резкого усиления процесса возбуждения в ЦНС, в частности, мотонейронов.
В связи с раскрытием ионных механизмов постсинаптического торможения появилась возможность и для объяснения механизма действия Вr. Натриq бромид в оптимальных дозах широко применяется в клинической практике как седативное (успокоительное) средство. Доказано, что такой эффект натрия бромида связан с усилением постсинаптического торможения в ЦНС. —
В центральной нервной системе постоянно функционируют два основных, взаимосвязанных процесса — возбуждение и торможение.
Торможение — это активный биологический процесс, направленный на ослабление, прекращение или предотвращение возникновения процесса возбуждения. Явление центрального торможения, т. е. торможения в ЦНС, было открыто И. М. Сеченовым в 1862 г. в опыте, получившим название «опыт сеченовского торможения». Суть опыта: у лягушки на срез зрительных бугров накладывали кристаллик поваренной соли, что приводило к увеличению времени двигательных рефлексов, т. е. к их торможению. Время рефлекса — это время от начала раздражения до начала ответной реакции.
Торможение в ЦНС выполняет две основные функции. Во-первых, оно координирует функции, т. е. оно направляет возбуждение по определенным путям к определенным нервным центрам, при этом выключая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата. Важность этой функции процесса торможения для функционирования организма можно наблюдать в эксперименте с введением животному стрихнина. Стрихнин блокирует тормозные синапсы в ЦНС (в основном глицинергические) и тем самым устраняет основу для формирования процесса торможения. В этих условиях раздражение животного вызывает некоординированную реакцию, в основе которой лежит диффузная (генерализованная) иррадиация возбуждения. При этом приспособителъная деятельность становится невозможной. Во-вторых, торможение выполняет охранительную или защитную функцию, пред охраняя нервные клетки от перевозбуждения и истощения при действии сверхсильных и длительных раздражителей.
Теории торможения . Н. Е. Введенским (1886) было показано, что очень частые раздражения нерва нервно-мышечного препарата вызывают сокращения мышцы в виде гладкого тетануса, амплитуда которого мала. Н. Е. Введенский полагал, что в нервно-мышечном препарате при частом раздражении возникает процесс пессимального торможения, т. е. торможение является как бы следствием перевозбуждения. Сейчас установлено, что его механизм заключается в длительной, застойной деполяризации мембраны, вызванной избытком медиатора (ацетилхолина), выделяющегося при частой стимуляции нерва. Мембрана полностью теряет возбудимость из-за инактивации натриевых каналов и не в состоянии ответить на приход новых возбуждений выделением новых порций медиатора. Таким образом, возбуждение переходит в противоположный процесс — торможение. Следовательно, возбуждение и торможение являются как бы одним и тем же процессом, возникают в одних и тех же структурах, с участием одного и того. же медиатора. Данная теория торможения называется унитарно-химической или монистической.
Медиаторы на постсинаптической мембране могут вызывать не только деполяризацию (ВПСП), но и гиперполяризацию (ТПСП). Эти медиаторы увеличивают проницаемость субсинаптической мембраны для ионов калия или хлора, в результате чего постсинаптическая мембрана гиперполяризуется и возникает ТПСП. Данная теория торможения получила название бинарно-химической, согласно которой торможение и возбуждение развиваются по разным механизмам, с участием тормозных и возбуждающих медиаторов соответственно.
Классификация центрального торможения. Торможение в ЦНС можно классифицировать по различным признакам:
По электрическому состоянию мембраны — деполяризационное и гиперполяризационное;
По отношению к синапсу — пресинаптическое и постсинаптическое;
По нейрональной организации — поступательное, латеральное (боковое), возвратное, реципрокное.
Постсинаптическое торможение развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется. Постсинаптическое торможение может быть деполяризационным, если в его основе лежит процесс длительной деполяризации, и гиперполяризационным, если — гиперполяризации.
Пресинаптическое торможение обусловлено наличием вставочных тормозных нейронов, которые формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению, например, к мотонейрону. В любом случае активации тормозного интернейрона, он вызывает деполяризацию мембраны афферентных терминалей, ухудшающей условия проведения по ним ПД, что таким образом уменьшает количество выделяемого ими медиатора, и, следовательно, эффективность синаптической передачи возбуждения к мотонейрону, что уменьшает его активность (рис. 14). Медиатором в таких аксо-аксональных синапсах является, по-видимому, ГАМК, которая вызывает повышение проницаемости мембраны для ионов хлора, которые выходят из терминали и частично, но длительно ее деполяризуют.
Рис. 14. Пресинаптическое торможение (схема): Н — нейрон, возбуждаемый афферентными импульсами, приходящими по волокну 1; Т — нейрон, образующий тормозные синапсы на пресинаптических разветвлениях волокна 1; 2 — афферентные волокна, вызывающие активность тормозного нейрона Т.
Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения (рис. 15).
Рис. 15. Схема поступательного торможения. Т — тормозньй нейрон
Возвратное торможение осуществляется вставочными тормозными нейронами (клетками Реншоу). Импульсы от мотонейронов, через отходящие от его аксона коллатерали, активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона (рис. 16). Это торможение реализуется за счет тормозных синапсов, образованных клеткой Реншоу на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, которая дает возможность стабилизировать частоту разряда мотонейрона и подавлять избыточную его активность.
Рис. 16. Схема возвратного торможения. Коллатерали аксона мотонейрона (1) контактируют с телом клетки Реншоу (2), короткий аксон которой, разветвляясь, образует тормозные синапсы на мотонейронах 1 и 3.
Латеральное (боковое) торможение. Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения (рис. 17). В таких случаях возбуждение направляется только по строго определенному пути.
Рис. 17. Схема латерального (бокового) торможения. Т — тормозный нейрон.
Именно латеральное торможение обеспечивает, в основном, системную (направленную) иррадиацию возбуждения в ЦНС.
Реципрокное торможение. Примером реципрокного торможения является торможение центров мышц-антагонистов. Суть этого вида торможения заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны (рис. 18). Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей.
Рис. 18. Схема реципрокного торможения. 1 — четырехглавая мышца бедра; 2 — мышечное веретено; 3 — сухожильный рецептор Гольджи; 4 — рецепторные клетки спиномозгового ганглия; 4а — нервная клетка, воспринимающая импульсы от мышечного веретена; 4б — нервная клетка, воспринимающая имульсы от рецептора Гольджи; 5 — мотонейроны, иннервирующие мышцы-разгибатели; 6 — тормозный промежуточный нейрон; 7 — возбуждающий промежуточный нейрон; 8 — мотонейроны, иннервирующие мышцы-сгибатели; 9 — мышца-сгибатель; 10 — моторные нервные окончания в мышцах; 11 — нервное волокно от сухожильного рецептора Гольджи.
Координированная работа антагонистических нервных центров обеспечивается формированием реципрокных отношений между нервными центрами благодаря наличию специальных тормозных нейронов – клеток Реншоу.
Известно, что сгибание и разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Сигнал от афферентного звена через промежуточный нейрон вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель, а через клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель (и наоборот).
Латеральное торможение
При латеральном торможении возбуждение, передаваемое через коллатерали аксона возбужденной нервной клетки, активирует вставочные тормозные нейроны, которые тормозят активность соседних нейронов, в которых возбуждение отсутствует или является более слабым.
В результате в этих соседних клетках развивается очень глубокое торможение. Образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону.
Латеральное торможение по нейронному механизму действия может иметь форму как постсинаптического, так и пресинаптического торможения. Играет важную роль при выделении признака в сенсорных системах, коре больших полушарий.
Значение торможения
Координация рефлекторных актов . Направляет возбуждение к определенным нервным центрам или по определенному пути, выключая те нейроны и пути, деятельность которых в данный момент является несущественной. Результатом такой координации является определенная приспособительная реакция.
Ограничение иррадиации .
Охранительное. Предохраняет нервные клетки от перевозбуждения и истощения. Особенно при действии сверхсильных и длительно действующих раздражителей.
В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности отдельных нервных клеток и нервных центров.
Координация – морфофункциональное взаимодействие нервных центров, направленное на осуществление определенного рефлекса или регуляции функции.
Морфологическая основа координации: связь между нервными центрами (конвергенция, дивергенция, циркуляция).
Функциональная основа: возбуждение и торможение.
Основные принципы координационного взаимодействия
Сопряженное (реципрокное) торможение .
Обратная связь .Положительная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в том же направлении, что и основные сигналы, что ведет к усилению рассогласования в системе.Отрицательная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в противоположном направлении и направлены на ликвидацию рассогласования, т.е. отклонений параметров от заданной программы (П.К. Анохин).
Общий конечный путь (принцип «воронки»Шеррингтона ). Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути».
Облегчение .Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов выше суммы реакций при изолированном раздражении этих рецептивных полей.
Окклюзия . Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов меньше, чем сумма реакций при изолированном раздражении каждого из рецептивных полей.
Доминанта .Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в ЦНС. ПоА.А. Ухтомскому , доминантный очаг характеризуется:
Повышенной возбудимостью,
Стойкостью и инертностью возбуждения,
Повышенной суммацией возбуждения.
Доминирующее значение такого очага определяет его угнетающее влияние на другие соседние очаги возбуждения. Принцип доминанты определяет формирование главенствующего возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
7. Субординация. Восходящие влияния преимущественно носят возбуждающий стимулирующий характер, нисходящие носят угнетающий тормозной характер. Эта схема согласуется с представлениями о росте в процессе эволюции роли и значении тормозных процессов в осуществлении сложных интегративных рефлекторных реакций. Имеет регулирующий характер.
Торможение в ЦНС — особый нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения.
Первичное постсинаптическое торможение — торможение, несвязанное с первоначальным процессом возбуждения и развивающееся в результате активации специальных тормозных структур. Тормозные синапсы образуют в своих окончаниях тормозной медиатор (ГАМК, глицин, в отдельных синапсах ЦНС роль тормозного медиатора может играть ацетилхолин). На постсинаптической мембране развивается тормозной постсинаптический потенциал (ТПСП), снижающий возбудимость мембраны постсинаптического нейрона. Тормозными нейронами могут служить только вставочные нейроны, афферентные нейроны всегда являются возбуждающими. В зависимости от вида тормозных нейронов и структурной организации нейронной сети постсинаптическое торможение подразделяется на:
- 1. Реципрокное торможение. Оно лежит в основе функционирования мышц-антагонистов и обеспечивает расслабление мышцы в момент сокращения мышцы-антагониста. Афферентное волокно, проводящее возбуждение от проприорецепторов мышц (например, сгибателей), в спинном мозге делится на две ветви: одна из них образует синапс на мотонейроне, иннервирующем мышцу-сгибатель, а другая — на вставочном, тормозном, образующем тормозной синапс на мотонейроне, иннервирующем мышцу-разгибатель. В результате возбуждение, приходящее по афферентному волокну, вызывает возбуждение мотонейрона, иннервирующего сгибатель и торможение мотонейрона мышцы-разгибателя.
- 2. Возвратное торможение. Оно реализуется через тормозные клетки Реншоу, открытые в спинном мозге. Аксоны мотонейронов передних рогов отдают коллатераль на тормозной нейрон Реншоу, аксоны которого возвращаются на тот же мотонейрон, образуя на нем тормозные синапсы. Таким образом формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов мотонейрона.
- 3. Центральное (Сеченовское) торможение. Оно осуществляется тормозными вставочными нейронами, через которые реализуется влияние на мотонейрон спинного мозга, возбуждения, возникающего в зрительных буграх под влиянием их раздражения. На мотонейроне спинного мозга суммируются ВПСП, возникающие в болевых рецепторах конечности и ТПСП, возникающие в тормозных нейронах под влиянием возбуждения таламуса и ретикулярной формации. В результате время защитного сгибательного рефлекса возрастает.
- 4. Латеральное торможение осуществляется с помощью тормозных вставочных нейронов в параллельных нейронных сетях.
- 5. Первичное пресинаптическое торможение развивается в терминальных отделах аксонов (перед пресинаптической структурой) под влиянием специальных аксо-аксональных тормозных синапсов. Медиатор этих синапсов вызывает деполяризацию мембраны терминалей и приводит их в состояние, подобное катодической депрессии Вериго. Мембрана в области такого бокового синапса препятствует проведению потенциалов действия к пресинаптической мембране, активность синапса уменьшается.
Пресинаптическим торможением называют снижение или выключение активности клетки за счет синаптического торможения оканчивающейся на ней возбуждающей терминали. Явление пресинаптического торможения зафиксировали Гассер и Грэхем в 1933 г., наблюдая эффект развития торможения сгибательных рефлексов при раздражении других корешков. Данный вид торможения термином “пресинаптическое торможение” впервые обозначили Фрэнк и Фуортес в 1957 г.
Увеличение частоты предварительных раздражений изменяет характер подавления. В частности, одна серия стимуляции с частотой 200-300 импульсов в секунду вызывает максимальное подавление менее чем на 10%, а две серии — подавление менее чем на 20%. При пресинаптическом торможении подавление моносинаптического ВПСП не связано с какими-либо изменениями их временных параметров.
Тормозные синапсы на окончаниях волокон обеспечивают довольно значительную деполяризацию, называемую деполяризацией первичных афферентов, или же первичную эфферентную деполяризацию (ПАД). В спинном мозгу ПАД обнаруживает длительную фазу (до 25 мс) нарастания до закругленной вершины и характеризуется большей продолжительностью по сравнению с постсинаптическими процессами. Большая продолжительность ПАД объясняется или длительным действием медиатора, или медленным, пассивным снижением деполяризации вследствие большой электрической постоянной времени мембраны. Пассивно снижающаяся компонента ПАД снимается импульсом, распространяющимся по афферентному волокну до его центральных окончаний.
Существует соответствие во всех отношениях между наблюдаемой деполяризацией первичных афферентных волокон и подавлением их синаптического возбуждающего действия.
Пресинаптическая деполяризация афферентов уменьшает величину их пресинаптического спайкового потенциала и таким образом уменьшает вызываемый им ВПСП. По данным Каца (1962), снижение спайкового потенциала на 5 мВ приводит к снижению выброса квантов медиатора и к снижению ВПСП до 50% и менее.
Характер ПАД в различных нейронах отличается по своим характеристикам. В целом временные параметры сравнимы. ПАД волокон кожного нерва отличается большей величиной амплитуды на одиночные раздражения с более коротким латентным периодом (около 2 мс), максимум также достигается ранее, чем в случае ПАД, вызываемых ритмическим раздражением нервных волокон, идущих от мышц. ПАД в клиновидном ядре имеет короткий латентный период (около 2 мс) и быстрый подъем до максимума.
Тормозные синапсы имеют химическую природу, медиатором в них служит ГАМК. Деполяризация первичных афферентов инактивирует возбуждающие натриевые каналы. Шунтирование натриевых каналов снижает амплитуду пресинаптического потенциала действия. В результате синаптическая передача моторного импульса ослабляется или исключается.
Во всех типах возбуждающих синапсов обнаруживается тесная зависимость между деполяризацией пресинаптических волокон и торможением синаптической передачи. Это торможение влияет не только на местные спинномозговые рефлексы, но также и на синаптические передачи в восходящих путях как от кожных афферентов, так и на спиноцеребеллярные. Кроме того, пресинаптическое торможение влияет на синаптические передачи задних столбов в ядра нежного и клиновидного пучков. Нисходящие импульсы из коры головного мозга и ствола мозга также оказывают пресинаптическое тормозное влияние на волокна группы и кожные афферентные волокна в спинном мозгу и клиновидном ядре. Обнаружено пресинаптическое торможение вторичных афферентных волокон отходящих от клиновидного ядра и имеющих переключение в таламусе. Синапсы с пресинаптическим торможением обнаружены в связанном с таламусом ядре мозга — латеральном коленчатом теле. В коре головного мозга не выявлено синаптических структур, которые могли бы осуществлять пресинаптическое торможение. На этих высших уровнях нервной системы доминирует постсинаптическое торможение. Пресинаптическое торможение действует как отрицательная обратная связь, уменьшая приток сенсорной информации в центральную нервную систему. Обычно эта отрицательная обратная связь не имеет точной топографии, но обычно концентрируется в пределах одной сенсорной модальности. Пресинаптическое торможение служит механизмом регуляции двигательных систем спинного мозга. Его особенностью является возможность специфического воздействия на отдельные синаптические входы без изменений возбудимости всей клетки. Таким образом, избыточная информация устраняется еще до того, как достигает места интеграции клеточного тела нейрона.
Вторичное торможение не связано с тормозными структурами, является следствием предшествующего возбуждения. Пессимальное торможение (открыто Н.Е. Введенским в 1886 г.) развивается в полисинаптических рефлекторных дугах при чрезмерной активации центральных нейронов и играет предохранительную роль. Оно выражается в стойкой деполяризации мембраны, приводящей к инактивации натриевых каналов. Торможение вслед за возбуждением» развивается в нейронах непосредственно после потенциала действия и характерно для клеток с длительной следовой гиперполяризацией. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов активности нейронов.
Механизмы координации рефлекторной деятельности: реципрокная иннервация, доминанта (А.А.Ухтомский), принципы обратной связи и общего конечного пути, принцип субординации.
Принцип иррадиации возбуждения. Иррадиация — распространение, расширение рефлекторного ответа. Это феномен “растекания” возбуждения по нейронам центральной нервной системы, развивающийся или после действия сверхсильного раздражителя, или на фоне выключения торможения. Распространение возбуждения возможно за счет многочисленных контактов между нейронами, возникающих при ветвлении аксонов и дендритов вставочных нейронов. Иррадиация позволяет увеличивать количество участвующих в рефлекторном ответе групп мышц. Ограничивают иррадиацию тормозные нейроны и синапсы.
На фоне действия стрихнина, блокирующего тормозные синапсы, наступают генерализованные судороги при тактильной стимуляции любого участка тела или при раздражении рецепторов любой сенсорной системы. В коре больших полушарий наблюдается явление иррадиации процесса торможения.
В основе координации рефлекторных актов лежат определенные механизмы, основанные на структурно-функциональной организации ЦНС и обозначаемые как “принципы” формирования рефлекторного ответа.
Принцип реципрокной иннервации. Реципрокная (сопряженная) координация открыта Н.Е. Введенским в 1896 году. Обусловлена реципрокным торможением, т.е. активация одного рефлекса одновременно сопровождается торможением второго, противоположного по своей физиологической сущности.
Принцип общего «конечного пути». Открыт английским физиологом Ч.Шеррингтоном (1906). Один и тот же рефлекс (например, сокращение мышцы) может быть вызван раздражением различных рецепторов, т.к. один и тот же конечный — мотонейрон передних рогов спинного мозга входит в состав многих рефлекторных дуг. Рефлексы, дуги которых имеют общий конечный путь, подразделяются на агонистические и антагонистические. Первые усиливают, вторые тормозят друг друга, как бы конкурируя за конечный результат. В основе подкрепления лежит конвергенция и суммация, в основе конкуренции за конечный путь — сопряженное торможение.
Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром. Обратная связь состоит во вторичной афферентации, поступающей в ЦНС от рецепторов, которые возбуждаются при изменении функциональной активности рабочего органа. Например, потенциалы действия, обусловленные возбуждением рецепторов мышц, сухожилий и суставных сумок сгибающейся конечности, в процессе осуществления акта сгибания поступают во все структуры ЦНС, начиная от центров спинного мозга. Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и отрицательную, когда рефлекс, ее вызывающий, тормозится. Обратная связь лежит в основе саморегуляции функций организма.
Принцип отдачи. Феномен отдачи состоит в быстрой смене одного рефлекса другим противоположного значения. Например, после сгибания конечности ее разгибание происходит быстрее, особенно если сгибание было сильным. Механизм этого явления состоит в том, что при сильном сокращении мышц возбуждаются рецепторы Гольджи сухожилий, которые через тормозные вставочные нейроны тормозят мотонейроны сгибательных мышц и образуют ветвь, которая возбуждает центр мышц — разгибателей. Благодаря этому механизму можно получить сумму рефлексов — цепные рефлексы (окончание одного рефлекторного ответа инициирует следующий) и ритмические (многократное повторение ритмичных движений).
Принцип доминанты. Конечный поведенческий эффект при координации рефлексов может быть изменен в зависимости от функционального состояния центров (наличия доминантных очагов возбуждения).
Особенности доминантного очага возбуждения:
- 1. Повышенная возбудимость нейронов.
- 2. Стойкость процесса возбуждения.
- 3. Способность к суммации возбуждения.
- 4. Иннертность. Очаг доминирует, подавляет соседние центры путем сопряженного торможения, возбуждаясь за их счет. Доминанту можно получить химическим воздействием на центры, например, стрихнином. В основе доминантного возбуждения лежит способность возбудительного процесса к иррадиации по нейронным цепям.
Физиология — наука, которая дает нам представление о человеческом организме и протекающих в нем процессах. Одним из таких процессов является торможение ЦНС. Оно представляет собой процесс, который порождается возбуждением и выражается в предупреждении появления другого возбуждения. Это способствует обеспечению нормальной деятельности всех органов и защищает нервную систему от перевозбуждения. Сегодня известно множество видов торможения, которые играют важную роль в работе организма. Среди них выделяют и реципрокное торможение(сочетанное), которое образуется в определенных тормозных клетках.
Виды центрального первичного торможения
Первичное торможение наблюдается в определенных клетках. Они находятся возле тормозных нейронов, которые производят нейротрансмиттеры. В ЦНС бывают такие виды торможения первичного: возвратное, реципрокное, латеральное торможение. Рассмотрим, как работает каждый из них:
- Латеральное торможение характеризуется затормаживанием нейронов тормозной клеткой, что находится около них. Часто этот процесс наблюдается между такими нейронами сетчатки глаз, как биполярные и ганглиозные. Это способствует созданию условий для отчетливого видения.
- Реципрокное — характеризуется взаимной реакцией, когда одни нервные клетки производят торможение других через вставочный нейрон.
- Возвратное — обуславливается торможением нейроном клетки, что тормозит этот же нейрон.
- Возвратное облегчение характеризуется понижением реакции иными тормозными клетками, при котором наблюдается уничтожение этого процесса.
В простых нейронах ЦНС происходит после возбуждения притормаживание, появляются следы гиперполяризации. Таким образом, реципрокное и возвратное торможение в происходят благодаря включению в цепь спинномозгового рефлекса особого тормозного нейрона, который именуется клеткой Реншоу.
Описание
В ЦНС постоянно работают два процесса — торможение и возбуждение. Торможение при этом направлено на прекращение или ослабление определенной деятельности в организме. Оно образуется при встрече двух возбуждений — тормозящего и тормозного. Реципрокное торможение представляет собой то, при котором возбуждение одних нервных клеток тормозит другие клетки через промежуточный нейрон, что имеет связь только с другими нейронами.
Экспериментальное открытие
Реципрокное торможение и возбуждение в ЦНС были выявлены и изучены Веденским Н.Е. Он проводил эксперимент на лягушке. На кожном покрове задней ее конечности осуществлялось возбуждение, которое вызывало сгиб и выпрямление конечности. Таким образом, согласованность этих двух механизмов представляет собой общую особенность всей нервной системы и наблюдается в головном и спинном мозге. Было установлено в ходе экспериментов, что совершение каждого действия движения основано на взаимосвязи торможения и возбуждения на одних и тех же нервных клетках ЦНС. Введенский Н.В говорил о том, что при возникновении возбуждения в какой-либо точке ЦНС вокруг этого очага появляется индукция.
Сочетанное торможение по Ч. Шеррингтону
Шеррингтон Ч. утверждает, что обеспечении полной согласованности конечностей и мышц. Этот процесс дает возможность конечностям сгибаться и выпрямляться. Когда человек сводит конечность, в колене образуется возбуждение, что переходит в спинной мозг на центр сгибательных мышц. Одновременно в центре разгибательных мышц появляется реакция замедления. Так происходит и наоборот. Запускается это явление при двигательных актах, имеющих большую сложность (прыжок, бег, ходьба). Когда человек идет, он поочередно сгибает и выпрямляет ноги. При согнутой правой ноге в центре сустава появляется возбуждение, в ином направлении происходит процесс торможения. Чем сложнее двигательные акты, тем большее число нейронов, которые несут ответственность за определенные мышечные группы, находятся в реципрокных отношениях. Таким образом, возникает благодаря работе вставочных нейронов спинного мозга, что отвечают за процесс торможения. Координированные отношения нейронов непостоянны. Изменчивость отношений между двигательными центрами дает возможность человеку делать непростые движения, например, играть на музыкальных инструментах, танцевать и прочее.
Реципрокное торможение: схема
Если рассматривать схематически этот механизм, то он имеет следующий вид: раздражитель, который поступает от афферентной части через обычный (вставочный) нейрон, вызывает возбуждение в нервной клетке. Нервная клетка приводит в движение мышцы-сгибатели, а через клетку Реншоу тормозит нейрон, что заставляет двигаться мышцы-разгибатели. Таким вот образом протекает координированное движение конечности.
Разгибание конечности происходит наоборот. Так, обеспечивает образование реципрокных отношений между центрами нервов определенных мышц благодаря клеткам Реншоу. Такое торможение является практичным с точки зрения физиологии, поскольку делает легким движение колена без какого-либо вспомогательного контролирования (произвольного или непроизвольного). Если бы этого механизма не было, то появилась бы механическая борьба мышц человека, судороги, а не скоординированные акты движения.
Суть сочетанного торможения
Реципрокное торможение позволяет организму делать произвольные движения конечностями: как легкие, так и достаточно сложные. Суть этого механизма заключается в том, что нервные центры противоположного действия находятся одновременно в противоположном состоянии. Например, при возбуждении центра вдоха центр выдоха заторможен. Если сосудосуживающий центр находится в возбужденном состоянии, то сосудорасширяющий в это время пребывает в заторможенном. Таким образом, сопряженное торможение центров рефлексов противоположного действия обеспечивает координацию движений и осуществляется с помощью специальных тормозных нервных клеток. Возникает согласованный сгибательный рефлекс.
Торможение по Вольпе
Вольпе в 1950 году было сформулировано предположение о том, что тревога представляет собой стереотип поведения, который закреплен в результате реакций на ситуации, которые ее вызывают. Связь между стимулом и реакцией может быть ослаблена в случае действия фактора, который тормозит тревогу, например, расслабление мышц. Вольпе назвал этот процесс «». Он лежит сегодня в основе метода поведенческой психотерапии — систематической десенситизации. В ее ходе пациента вводят во множество представляемых ситуаций, одновременно вызывается мышечное расслабление при помощи транквилизаторов или гипноза, которое снижает уровень тревоги. По мере закрепления отсутствия тревоги в легких ситуациях, пациент переходит к сложным ситуациям. В результате терапии человек приобретает навыки самостоятельно контролировать тревожные ситуации в реальности при помощи техники мышечного расслабления, которой он овладел.
Таким образом, реципрокное торможение было открыто Вольпе и широко применяется сегодня в психотерапии. Суть метода заключается в том, что происходит уменьшение силы определенной реакции под воздействием иной, которая была вызвана одновременно. Этот принцип находится в основе конт-обуславливания. Сочетанное торможение обуславливается тем, что реакция страха или тревоги затормаживается эмоциональной реакцией, которая возникает одновременно и является несовместимой со страхом. Если такое торможение происходит периодически, то условная связь между ситуацией и реакцией тревоги ослабевает.
Метод психотерапии Вольпе
Джозеф Вольпе обратил внимание на то, что привычкам свойственно угасать в случае развития новых привычек в одинаковой ситуации. Он использовал термин «реципрокное торможение» для описания ситуаций, где появление новых реакций приводит к угасанию ранее возникавших реакций. Так, при одновременном присутствии стимулов к появлению несовместимых реакций, развитие доминирующей реакции в определенной ситуации предполагает сопряженное торможение других. На основании этого он разработал метод лечения тревожности и страхов у людей. Этот способ предполагает нахождение тех реакций, что подходят для возникновения реципрокного торможения реакций страха.
Вольпе выделял следующие реакции, что несовместимы с тревогой, применение которых даст возможность изменить поведение человека: реакции ассертивные, сексуальные, релаксация и «облегчение тревоги», а также дыхательные, моторные, медикаментозно усиленные реакции и те, что вызваны беседой. На основании всего этого были разработаны различные техники и приемы в психотерапии при лечении тревожных пациентов.
Итоги
Таким образом, на сегодняшний день учеными объяснен рефлекторный механизм, который использует реципрокное торможение. Согласно этому механизму нервные клетки возбуждают тормозные нейроны, которые находятся в спинном мозге. Это все способствует координированному движению конечностей у человека. Человек имеет способность совершать различные сложные двигательные акты.
нервной системы человека | Описание, развитие, анатомия и функции
Пренатальное и постнатальное развитие нервной системы человека
Почти все нервные клетки или нейроны генерируются во время пренатальной жизни, и в большинстве случаев после этого они не заменяются новыми нейронами. Морфологически нервная система впервые появляется примерно через 18 дней после зачатия в результате образования нервной пластинки. Функционально он появляется с первым признаком рефлекторной активности во втором пренатальном месяце, когда стимуляция прикосновением к верхней губе вызывает реакцию отдергивания головы.Многие рефлексы головы, туловища и конечностей могут появиться на третьем месяце.
В процессе своего развития нервная система претерпевает значительные изменения, чтобы достичь своей сложной организации. Чтобы произвести примерно 1 триллион нейронов, присутствующих в зрелом мозге, в среднем в течение всей пренатальной жизни необходимо генерировать 2,5 миллиона нейронов в минуту. Это включает формирование нейронных цепей, содержащих 100 триллионов синапсов, поскольку каждый потенциальный нейрон в конечном итоге связан либо с выбранным набором других нейронов, либо с конкретными целями, такими как сенсорные окончания.Более того, синаптические связи с другими нейронами устанавливаются в определенных местах на клеточных мембранах целевых нейронов. Совокупность этих событий не считается исключительным продуктом генетического кода, поскольку генов просто не хватает, чтобы объяснить такую сложность. Скорее, дифференцировка и последующее развитие эмбриональных клеток в зрелые нейроны и глиальные клетки достигается двумя наборами влияний: (1) специфическими подмножествами генов и (2) стимулами окружающей среды внутри и вне эмбриона.Генетические влияния имеют решающее значение для развития нервной системы в упорядоченной и временной последовательности. Клеточная дифференцировка, например, зависит от серии сигналов, регулирующих транскрипцию, процесса, в котором молекулы дезоксирибонуклеиновой кислоты (ДНК) дают начало молекулам рибонуклеиновой кислоты (РНК), которые, в свою очередь, выражают генетические сообщения, контролирующие клеточную активность. Влияния окружающей среды, происходящие от самого эмбриона, включают клеточные сигналы, которые состоят из диффундирующих молекулярных факторов ( см. Ниже Развитие нейронов).К факторам внешней среды относятся питание, сенсорный опыт, социальное взаимодействие и даже обучение. Все это важно для правильной дифференциации отдельных нейронов и тонкой настройки синаптических связей. Таким образом, нервная система требует непрерывной стимуляции на протяжении всей жизни для поддержания функциональной активности.
Развитие нейронов
На второй неделе пренатальной жизни быстро растущая бластоциста (связка клеток, на которую делится оплодотворенная яйцеклетка) превращается в так называемый эмбриональный диск.Эмбриональный диск вскоре приобретает три слоя: эктодерму (внешний слой), мезодерму (средний слой) и энтодерму (внутренний слой). Внутри мезодермы растет хорда, осевой стержень, который служит временным позвоночником. И мезодерма, и хорда выделяют химическое вещество, которое заставляет соседние недифференцированные клетки эктодермы утолщаться вдоль того, что станет дорсальной средней линией тела, образуя нервную пластинку. Нервная пластинка состоит из нервных клеток-предшественников, известных как нейроэпителиальные клетки, которые развиваются в нервную трубку ( см. Ниже Морфологическое развитие).Затем нейроэпителиальные клетки начинают делиться, диверсифицироваться и давать незрелые нейроны и нейроглию, которые, в свою очередь, мигрируют из нервной трубки в свое окончательное местоположение. Каждый нейрон образует дендриты и аксон; аксоны удлиняются и образуют ветви, концы которых образуют синаптические связи с выбранным набором целевых нейронов или мышечных волокон.
Человеческое эмбриональное развитиеРазвитие человеческого эмбриона на 18-й день, на стадии диска или щита, показано на трех четвертях (слева) и в поперечном сечении (справа).
Британская энциклопедия, Inc. Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасЗамечательные события этого раннего развития включают упорядоченную миграцию миллиардов нейронов, рост их аксонов (многие из которых широко распространяются по всему мозгу) и формирование тысяч синапсов между отдельными аксонами и их целевыми нейронами. Миграция и рост нейронов зависят, по крайней мере частично, от химических и физических воздействий.Растущие концы аксонов (называемые конусами роста), по-видимому, распознают и реагируют на различные молекулярные сигналы, которые направляют аксоны и нервные ветви к их соответствующим целям и устраняют те, которые пытаются синапсировать с неподходящими целями. Как только синаптическая связь установлена, клетка-мишень высвобождает трофический фактор (например, фактор роста нервов), который необходим для выживания нейрона, синапсирующегося с ней. Сигналы физического наведения участвуют в наведении контактов или миграции незрелых нейронов по каркасу из глиальных волокон.
В некоторых регионах развивающейся нервной системы синаптические контакты изначально не являются точными или стабильными, и позже за ними следует упорядоченная реорганизация, включая устранение многих клеток и синапсов. Нестабильность некоторых синаптических связей сохраняется до наступления так называемого критического периода, до которого влияние окружающей среды играет значительную роль в правильной дифференцировке нейронов и в тонкой настройке многих синаптических связей. После критического периода синаптические связи становятся стабильными и вряд ли будут изменены под влиянием окружающей среды.Это говорит о том, что на определенные навыки и сенсорную деятельность можно повлиять во время развития (включая послеродовую жизнь), а для некоторых интеллектуальных навыков эта способность к адаптации предположительно сохраняется во взрослой жизни и в конце жизни.
Что такое взаимность?
Что такое взаимность?
Взаимность — это процесс обмена вещами с другими людьми для получения взаимной выгоды. Норма взаимности, иногда называемая правилом взаимности, является социальной нормой, когда, если кто-то что-то делает для вас, вы чувствуете себя обязанным ответить на услугу.Взаимодействие с другими людьми
Процесс социализации играет важную роль в развитии этой потребности отвечать взаимностью. Благодаря опыту дети учатся делиться с другими, по очереди и взаимодействовать друг с другом. Взаимность играет важную роль в развитии и продолжении отношений. Он также играет важную роль в убеждении или побуждении других принять определенные убеждения или поведение.
Как узнать?
Вы когда-нибудь чувствовали себя обязанными что-то сделать для кого-то, потому что он сначала что-то сделал для вас? Норма взаимности — это всего лишь один из типов социальных норм, который может оказать сильное влияние на наше поведение.
Норма взаимности работает по простому принципу: люди склонны чувствовать себя обязанными отвечать за услугу после того, как люди оказали им услугу.
Когда ваши новые соседи приносят тарелку печенья, чтобы поприветствовать вас по соседству, вы можете почувствовать себя обязанным ответить вам одолжением, когда они попросят вас позаботиться об их собаке, пока они в отпуске.
Типы взаимности
Существует три основных типа взаимности:
- Обобщенная взаимность : Эта форма часто подразумевает обмены внутри семьи или друзей.Нет ожидания ответной услуги; вместо этого люди просто делают что-то для другого человека, исходя из предположения, что этот человек сделает то же самое для них. Этот тип взаимности связан с альтруизмом.
- Сбалансированная взаимность : Этот тип включает расчет стоимости обмена и ожидание того, что услуга будет возвращена в течение определенного периода времени. Например, кто-то может обменять что-то, что у него есть, будь то умение или материальный предмет, на что-то представляющее равную ценность.
- Отрицательная взаимность : эта форма взаимности возникает, когда одна сторона, участвующая в обмене, пытается получить от нее больше, чем другая сторона. Продажа столь необходимого предмета по завышенной цене — один из примеров отрицательной взаимности.
использует
Одна из областей, где эта норма обычно применяется, — это маркетинг. Маркетологи используют широкий спектр стратегий, чтобы убедить потребителей совершить покупки. Некоторые из них просты, например распродажи, купоны и специальные акции.Другие гораздо более тонкие и используют принципы человеческой психологии, о которых многие даже не подозревают.
Другие примеры взаимности включают:
- Продавец дает халяву потенциальному покупателю в надежде, что это заставит его вернуть услугу, купив что-то
- Лидер, предлагающий своим последователям внимание и наставничество в обмен на лояльность
- Предлагаем клиентам ценную информацию в обмен на подписку на будущие маркетинговые предложения
Влияние взаимности
У взаимности есть несколько очевидных преимуществ.Во-первых, забота о других помогает выживанию вида.
В ответ мы гарантируем, что другие люди получат помощь, когда они в ней нуждаются, и что мы получим помощь, когда она нам понадобится.
Взаимность также позволяет людям делать то, что они не смогли бы сделать в одиночку. Работая вместе или обмениваясь услугами, люди могут достичь большего, чем по отдельности.
Взаимность и убеждение
Существует также ряд приемов убеждения, использующих тактику взаимности.Эти стратегии используются людьми, которые пытаются убедить вас действовать или выполнить просьбу, например продавцами или политиками.
Один из них известен как метод «это еще не все». Допустим, вы покупаете новый мобильный телефон. Продавец показывает ваш телефон и называет цену, но вы все еще не совсем уверены. Если продавец предлагает добавить чехол для телефона без дополнительной оплаты, вы можете подумать, что он делает вам одолжение, что, в свою очередь, может заставить вас почувствовать себя обязанным купить телефон.
Советы по навигации по взаимности
Во многих случаях норма взаимности — это действительно хорошо. Это помогает людям вести себя социально приемлемым образом и позволяет им соглашаться друг с другом. Но что делать, если вы пытаетесь преодолеть желание ответить взаимностью, например, пытаетесь избежать необходимости покупать предмет после получения халявы?
Несколько советов, которые могут помочь:
- Подождите немного. Эксперты предполагают, что желание ответить взаимностью наиболее сильно проявляется сразу после первого обмена.Если вы можете подождать, вы, вероятно, почувствуете меньшее давление, чтобы вернуть услугу.
- Оценить обмен. Подумайте, соответствует ли услуга ожидаемой прибыли. Во многих случаях первоначальный подарок или услуга намного меньше запрашиваемой ответной услуги.
Понимание того, как норма взаимности влияет на поведение, может помочь вам лучше оценивать убедительные сообщения и просьбы.
Возможные ловушки
Взаимность не всегда является равномерным обменом, что открывает возможности для дисбаланса или даже злоупотреблений.Исследования показали, что люди часто готовы оказать пропорционально большую услугу после того, как кто-то сделал для них что-то незначительное.
Участие в этом первом взаимном обмене может повысить вероятность того, что вы ответите на другие, часто более крупные, запросы в будущем. В маркетинге это часто называют техникой «ступни в дверь». Кто-то начинает с небольшого запроса, а как только вы соглашаетесь с ним, он отправляет гораздо больший запрос.
Другой подход, известный как метод «дверь в лицо», также может быть использован для использования принципа взаимности.Убежденные начинают с того, что просят об очень большой услуге, которую, как они знают, вы откажетесь. Затем кажется, что они уступают, прося об одолжении гораздо меньшего, которое вы можете почувствовать своим долгом.
На самом деле целью все время была небольшая услуга, но, оказав вам услугу, сделав меньшую просьбу, вы чувствуете себя обязанным ответить на просьбу, сказав «да» на меньшую просьбу.
История взаимности
Один плодотворный эксперимент показал, насколько сильной может быть взаимность в реальном мире.В 1974 году социолог Филипп Кунц провел эксперимент: он разослал рукописные рождественские открытки с запиской и фотографией его семьи и его семьи примерно 600 случайно выбранным людям. Все получатели карточек были совершенно незнакомыми людьми. Вскоре после отправки открыток начали поступать отклики.
Кунц получил почти 200 ответов. Почему так много людей ответят совершенно незнакомому человеку? Это правило взаимности в действии. Поскольку Кунц что-то для них сделал (отправил вдумчивую записку во время курортного сезона), многие получатели чувствовали себя обязанными вернуть услугу.
Взаимное торможение и как оно применимо к вам
Термин взаимное торможение может не иметь большого значения для вас, но есть несколько вещей, которые влияют на ваше тело более регулярно. Теория реципрокного торможения гласит: «Когда центральная нервная система посылает агонисту (мышцу, вызывающую движение) сообщение о сокращении, напряжение в антагонисте (мышечное противодействующее движение) подавляется импульсами от мотонейронов, и поэтому она должна одновременно расслабляться. », Взято из« Принципов и практик массажа »Сьюзан Сальво.Это означает, что наши мышцы действуют парами и координируются друг с другом, одновременно расслабляясь и сокращаясь в качестве защитной меры, чтобы уберечь нас от травм. Если обе противостоящие мышцы задействуют одновременно, не позволяя другой расслабиться, может произойти разрыв мышцы. Типичный пример этого — бег. Удар по земле будет посылать импульсы от центральной нервной системы для сокращения и расслабления противоположных мышц (подколенных сухожилий и четырехглавой мышцы бедра) для обеспечения плавного и безопасного движения.
Хотя большая часть взаимного торможения контролируется подсознательно, мы можем использовать этот принцип, чтобы «обмануть» тело во время растяжки, чтобы добиться большего диапазона движений. Например, если цель состоит в том, чтобы растянуть подколенные сухожилия, сокращение четырехглавой мышцы при достижении ближнего конечного диапазона движения позволит подколенным сухожилиям еще больше расслабиться, тем самым увеличивая растяжение. Другой пример — во время растяжки мышц груди (большая / малая грудная мышца). Достигнув конечного диапазона движения этой растяжки, сократите мышцы, расположенные непосредственно за плечом (задние дельтовидные / средние трапециевидные), чтобы отправить сигнал вашему телу, заставляющий мышцы груди еще больше расслабиться.
Растяжка должна быть неотъемлемой частью тренировки, однако простая растяжка — не самый эффективный способ обеспечить надлежащий мышечный баланс в вашем теле. Несмотря на то, что существует множество различных форм растяжки, которые могут резко повлиять на увеличение диапазона движений, без устранения дисбаланса силы и напряжения противоположных мышц, эти результаты часто носят временный характер. Если цель состоит в том, чтобы растянуть мышцы груди, чтобы исправить наклон плеча вперед, очень важно также укрепить мышцы спины, которые отвечают за удержание плеч на месте.Чтобы исправить чрезмерный наклон таза вперед, важно не только растянуть сгибатели бедра, но и укрепить ягодицы. Укрепляя эти противоположные мышцы, вы ослабите напряжение, создаваемое слишком напряженными мышцами, позволяя диапазону движений, полученному при растяжке, иметь длительный эффект. Если вы хотите узнать больше о том, как правильно сбалансировать свои тренировки или получить дополнительную информацию о том, как эффективно растягиваться, обратитесь к личному тренеру по фитнесу Уиллу Пэтону.
Консультации по фитнесу, спортивная подготовка, силовые тренировки
здание, клуб, фитнес, здоровье, мышца, персональный тренер, персональный тренинг, сиэтл, сила, растяжка, поднятие тяжестей, тренировка
Reciprocal Innervation — обзор
Сложность управления 12 экстраокулярными мышцами значительно снижается несколькими «законами», которые уменьшают степень свободы управления глазами. Эти законы реализуются в глазодвигательной анатомии около выходного каскада, и поэтому почти все движения глаз подчиняются им.
III.E.2 Закон моторного соответствия Геринга
Закон Геринга уменьшает вдвое степени свободы глазодвигательной системы с 6 до 3, поскольку он утверждает, что два глаза действуют в унисон (сопряженно) для всех движений глаз, кроме вергентности. Таким образом, мозг работает так, как будто есть только один глаз, циклопическая сетчатка. Это моторное соответствие достигается за счет соединения пар мышц двух глаз. Например, латеральная прямая мышца правого глаза и медиальная прямая мышца левого глаза составляют пару коромысел.Они получают одинаковые команды и совершают движение вправо.
Цепи ствола головного мозга для реципрокной иннервации и моторного соответствия при горизонтальных движениях глаз (медиальная и латеральная прямая мышца) показаны на рисунке 5 для быстрых движений глаз, таких как саккады, и на рисунке 6 для плавных / медленных движений глаз. такие как VOR. Привязка к горизонтальным движениям проста: сигналы, поступающие в правое отводящее ядро, не только иннервируют мотонейроны боковой прямой мышцы правого глаза, но также иннервируют интернейроны в отводящем ядре.Эти интернейроны передают этот сигнал через медиальный продольный пучок в контрлатеральное глазодвигательное ядро, синапсируя с OMN медиальной прямой мышцы левого глаза. Для реципрокной иннервации тормозящая копия командного сигнала направляется к контралатеральной отводящей мышце, которая расслабляет контралатеральную латеральную прямую мышцу, а также посредством цепи яркости расслабляет ипсилатеральную медиальную прямую мышцу.
Рис. 5. Нейронная цепь для сопряженного движения глаза вправо. Сопряженное движение глаз, как на этом рисунке, вызывает активность в схематической нейронной цепи, которая реализует как закон Геринга (двигательное соответствие), так и закон Декарта-Шеррингтона (реципрокная иннервация).При движении глаза вправо сжатые мышцы-агонисты представляют собой правую латеральную прямую мышцу и левую медиальную прямую мышцу, а расслабленные / подавленные мышцы-антагонисты представляют собой левую боковую прямую мышцу и правую медиальную прямую мышцу. В показанном примере сигнал для совершения правой саккады исходит из левого полушария (кора и холмик) и передается через нейроны возбуждающего импульса (EBN) к мотонейронам правой боковой прямой мышцы живота в n. VI (похищает). Чтобы также сократить скрученную мышцу в левом глазу (медиальную прямую мышцу) и тем самым сделать движение сопряженным, набор немоторных нейронов в n.VI передают сигнал движения контралатеральному n. III (глазодвигательный). Взаимная иннервация (ингибирование двух мышц-антагонистов) осуществляется ингибирующими взрывными нейронами (IBN), которые проецируются влево n. VI и, таким образом, оба напрямую ингибируют мотонейроны левой боковой прямой мышцы живота и косвенно уменьшают сокращение правой медиальной прямой мышцы живота через n. VI до н. III проекция. Подобные схемы обеспечивают моторное соответствие и реципрокную иннервацию для других экстраокулярных мышц, чтобы выполнять сопряженные вертикальные и торсионные движения.
Рис. 6. Нервная цепь горизонтального вестибулоокулярного рефлекса (VOR). На диаграмме изображены горизонтальные движения глаз вправо, которые компенсируют поворот головы влево. Показанная здесь базовая возможность подключения обслуживает все сопряженные движения глаза вправо (например, рис. 5), независимо от источника сигнала команды движения глаз. Для VOR этот сигнал исходит из полукружных каналов и передается через биполярные нейроны вестибулярного ганглия (VG) к вестибулярным ядрам (VN).VN посылает возбуждающую проекцию на контралатеральные глазодвигательные нейроны в n. VI (abducens), а также направляет тормозную проекцию на ипсилатеральную n. VI. Последствия повышенной активности левого горизонтального канала усиливаются снижением активности правого горизонтального канала, иллюстрируя двухтактную работу пар каналов. Дополнительный возбуждающий путь (не показан) от VN непосредственно к медиальным мотонейронам прямой мышцы живота в ипсилатеральном n. III (глазодвигательный) дает медиальной прямой мышце свой VOR-привод с тремя нейронами в дополнение к VOR-возбуждению с четырьмя нейронами через контралатеральный n.VI.
Для остальных восьми мышц запирание менее очевидно. Каждая верхняя косая мышца и противоположная нижняя прямая мышца составляют одну пару коромысел, а каждая нижняя косая мышца и противоположная верхняя прямая мышца — другую. Эти пары отражают истинные оси вращения этих мышц и их отношение к вестибулярным каналам (Рис. 7)
Рис. 7. Оси вращения глазных мышц и каналов. (Вверху, слева и справа) Ось вращения для всех экстраокулярных мышц, кроме медиальной и латеральной прямой мышцы живота (внизу, слева и справа).Для каждой мышцы направление или знак вращения глаза задается правилом левой руки: направьте большой палец вдоль стрелки, и изгиб (слегка сомкнутых) пальцев указывает на вращение глаза. Оси вращения шести полукружных каналов (внизу в середине) перпендикулярны плоскостям каналов, изображенным выше (вверху в середине), и показывают (по правилу левой руки) вращение глаза, вызванное стимуляцией отдельных каналов. Легко увидеть, какая ось мышц каждого глаза лучше всего совпадает с осью каждого канала; эти мышцы получают возбуждающую проекцию из трех нейронов из канала.Хотя у приматов глаза переместились с боковой стороны головы вперед, каналы мало изменились, а экстраокулярные мышцы приматов сохранили свои оси вращения в соответствии с осями каналов. На рисунке также графически показано обоснование соединения нижней косой мышцы с контралатеральной верхней прямой мышцей и верхней косой мышцы с контралатеральной нижней прямой мышцей. Угловая разница в обоих случаях намного меньше, чем разница между осями левой и правой косых мышц или левой и правой осями верхней и нижней прямой мышцы живота.Этот рисунок был вдохновлен рис. 51 у Геринга, 1942. Значения его оси, которые использовались здесь, имеют ось верхнего-нижнего прямых мышц, находящуюся на 29 ° от оси для чисто вертикальных движений и ось верхнего-нижнего наклонные отклонения от оси крутильного движения на 38 °. Параллелограмм [a, a ′, a ″] (вверху слева) — это его геометрическое построение чистого подъемного движения (без скручивания) из одновременных сокращений нижней косой и верхней прямой мышцы живота: сложите векторы осей вращения этих двух мышц. чтобы получить ось для комбинированного вращения.
III.E.3 Законы листинга и Дондерса и потеря кручения
У приматов крутильные движения сильно ограничены. Например, человеческий глаз поворачивается только на несколько градусов в ответ на большой наклон головы вбок (технически «крен»), в то время как глаз кролика поворачивается до 70 °. Отсутствие значительного крутильного движения имеет эволюционное преимущество для стереоскопического зрения.
Скручивание глаза классически описывается законами Дондерса и Листинга. Закон Дондерса утверждает, что каждое направление взгляда (комбинация определенного азимута и определенного возвышения) всегда связано с уникальным скручиванием, независимо от последовательности движений глаз, используемых для достижения этого конкретного азимута / возвышения.Это имеет важные последствия. Во-первых, хотя для определения общего вращения требуются три координаты, закон Дондерса означает, что глаз имеет только 2 степени свободы, и, следовательно, необходимо указать только два угла (азимут и угол места). Это аналогично навигации; долготы и широты достаточно, даже если места на поверхности океана имеют три координаты в пространстве. Во-вторых, в то время как трехмерное вращение не является коммутативным, закон Дондерса предусматривает, что движения человеческого глаза фактически коммутируют.
Закон о листинге подтверждает закон Дондерса и многое другое, потому что он определяет конкретное кручение, связанное с любым конечным направлением взгляда, тогда как закон Дондерса только гарантирует, что это кручение является уникальным. Чтобы найти положение скручивания, которое глаз всегда будет принимать при определенном азимуте и возвышении, закон Листинга предписывает найти ось во фронтальной плоскости через центр глаза (плоскость Листинга), так что одно вращение вокруг этой оси захватывает глаз от основного положения к конкретному рассматриваемому азимуту и углу места.Это единственное вращение позволит получить ориентацию, которую глаз всегда будет иметь при этом азимуте и возвышении, независимо от того, какая последовательность движений глаза фактически использовалась, чтобы попасть туда. Это означает, что человеческий глаз имеет нулевое скручивание после чисто горизонтальных или чисто вертикальных движений из основного положения (это называется «второстепенными» положениями). Однако для типичного наклонного направления («третичное» положение глаза) глаз, кажется, претерпел крутильное вращение, потому что в реальном мире вертикальные линии больше не проходят вдоль вертикального меридиана сетчатки при фиксации.Это часто называют «ложным скручиванием», потому что глазу не нужно было совершать истинное крутильное движение вокруг луча зрения, а, скорее, только один поворот вокруг оси в плоскости Листинга, которая ортогональна оси кручения.
Экстраокулярные мышцы играют сложную роль в контроле скручивания глаза. Медиальная и латеральная прямая мышцы живота имеют минимальные торсионные движения; однако, как отмечалось ранее, четыре оставшихся мышцы (нижняя косая, верхняя косая, верхняя прямая мышца и нижняя прямая мышца) имеют как вертикальное, так и крутильное действие («смешанные» действия).Следовательно, как сказал Эвальд Геринг, подъем взгляда осуществляется «только за счет взаимодействия верхней прямой мышцы живота и нижней косой мышцы живота». Точно так же депрессия вызывается «взаимодействием нижней прямой мышцы живота с верхней косой мышцей». Это аналогично поднятию штанги: если обе руки поднимаются с одинаковой силой, штанга будет оставаться ровной, тогда как если одна рука прилагает слишком много силы, штанга будет наклоняться (искажаться) при подъеме. Точно так же, когда верхняя косая мышца и нижняя прямая мышца сокращаются совместно, их направленные вниз силы складываются, но выталкивающая сила от повышенного напряжения в верхней косой мышце компенсируется внутренней силой от повышенного напряжения в нижней косой мышце.Более того, повышенное напряжение верхней косой и нижней прямой мышцы живота всегда компенсируется уменьшением напряжения нижней косой и верхней прямой мышцы живота соответственно. Это приводит к уменьшению направленной вверх силы, но с уменьшением выталкивания и отмены вторжения. Таким образом, глаза могут двигаться вниз (или вверх) с очень небольшим скручиванием, даже если все четыре задействованные мышцы имеют значительные скручивающие действия.
Законы Дондерса и Листинга не являются абсолютными и неотъемлемыми от пассивных свойств глаза.Большие отклонения сопровождают конвергенцию, наклон головы, сон и некоторые невральные патологии. Торсионный нистагм, явное нарушение законов Дондерса и Листинга, может быть получен с помощью электростимуляции определенных ядер среднего мозга или отдельных каналов, а также может быть следствием центрального заболевания.
Реципрокное ингибирование — обзор
55.4 Созревание ГАМК
A и хлоридных каналов, управляемых рецепторами глицинаИ ГАМК, и глицин вносят вклад в реципрокное ингибирование, лежащее в основе чередования левого и правого каналов на E18.5, в то время как ГАМК имеет небольшой вклад, если таковой имеется, в это чередование после рождения, предполагая, что относительный вклад ГАМК А (ГАМК А R) и рецепторов глицина (GlyRs) изменяется во время развития (Рисунок 55.2 (а); Nakayama et al., 2002; см. Также Hinckley et al., 2005; Pflieger et al., 2002). Ziskind-Conhaim и его коллеги исследовали свойства миниатюрных тормозных постсинаптических токов (mIPSC) в поясничных МН крыс во время перинатального развития (E17-18 — P1-3; Gao and Ziskind-Conhaim, 1995; Gao et al., 2001). В первую очередь, миниатюрные mIPSC большой продолжительности регистрируются в эмбриональных MNs, тогда как mIPSC меньшей продолжительности становятся очевидными после рождения. Укорочение mIPSC связано с увеличением как частоты, так и амплитуды. Нанесение в ванну антагонистов GABA A R или GlyR (бикукуллин и стрихнин, соответственно) выявляет два типа фармакологически различных mIPSC: GlyR-опосредованные с большой амплитудой / быстро распадающиеся и опосредованные GlyR с малой амплитудой / медленно распадающиеся GABA A R-опосредованные. МПСК.Средняя амплитуда глицинергических mIPSC увеличивается вдвое после рождения, в то время как нет значительного изменения амплитуды чистого ГАМКергического и смешанного тока глицин-ГАМК. Частота GABA A R-опосредованных mIPSCs на E17-18 намного выше, чем частота GlyR-опосредованных mIPSCs, подтверждая, что GABA является функционально доминирующей на ранних стадиях. Наиболее ярким изменением в развитии является восьмикратное послеродовое увеличение отношения частоты mIPSC, опосредованных глицином и ГАМК, что указывает на переход от ГАМК к глицину во время перинатального развития (Рисунок 55.2 (б); Гао и др., 2001). Эти результаты объясняют связанный с возрастом переход от ГАМК к глицину во вкладе ингибирующих аминокислот в реципрокное лево-правое торможение во время передвижения.
Рисунок 55.2. Связанные с возрастом изменения относительного вклада ГАМК , и глициновых рецепторов в ингибирование синаптической передачи. (а) Влияние применения 10 мкМ бикукуллина (средняя панель) или 5 мкМ стрихнина (правая панель) на двигательный паттерн, индуцированный 20 мкМ серотонином на E18.5 (верхние трассы) и E20.5 (нижние трассы). Лево-правое чередование основывается как на GABA , A , так и на рецепторах глицина на E18.5 и только на рецепторах глицина на E20.5. Перерисовано с разрешения Накаямы К., Нисимару Х. и Кудо Н. (2002). Основа изменений лево-правой координации ритмической двигательной активности во время развития спинного мозга крысы. Журнал неврологии 22: 10388–10398. (b) Чистая ГАМК A рецептор-опосредованные миниатюрные ингибирующие постсинаптические токи (mIPSCs), зарегистрированные в поясничном отделе спинного мозга крысы in vitro , составляют около половины популяции mIPSC на E17-18 и уменьшаются на P1-3.Напротив, вклад mIPSCs, опосредованных чистым глициновым рецептором (GlyR), увеличивается в перинатальный период. Доля смешанных GlyR-GABA A R-обеспечиваемых mIPSCs остается относительно постоянной (около 20-25%). Перерисовано с разрешения Gao BX, Stricker C и Ziskind-Conhaim L (2001). Переход от ГАМКергической к глицинергической синаптической передаче во вновь сформированных спинномозговых сетях. Журнал нейрофизиологии 86: 492–502. (c) Доля ГАМК, глицина и смешанных явлений в конце первой постнатальной недели у контрольных животных и после неонатальной перерезки спинного мозга (SCT).Этот SCT предотвращает подавление онтогенетической регуляции mIPSCs GABA A R, тогда как повышенная регуляция GlyR затрагивается незначительно.
Перерисовано с разрешения Садлауда К., Тазерарта С., Брокарда С. и др. (2010) Дифференциальная пластичность ГАМКергической и глицинергической синаптической передачи к поясничным мотонейронам крысы после повреждения спинного мозга. Журнал неврологии 30: 3358–3369.В поддержку этих электрофизиологических данных увеличение количества глицинсодержащих терминалов, контактирующих с MNs, более важно, чем увеличение количества GABA-содержащих терминалов (Simon and Horcholle-Bossavit, 1999).Кроме того, плотность GlyR увеличивается с P1 до P7, тогда как плотность субъединиц GABA A R падает в тот же период (Sadlaoud et al., 2010). В этом исследовании в день рождения была проведена ТСК для изучения вклада ствола мозга в переход от ГАМК к глицину в процессе развития. У животных P7 с таким неонатальным пересечением плотности GlyR не изменились по сравнению с контролем того же возраста, в то время как подавление развития GABA A R было предотвращено.Запись цельноклеточных патч-кламп-регистрации mIPSC, выполненная из поясничных MNs на P7, показала, что постоянная времени распада миниатюрных IPSC и доля ГАМКергических событий были значительно увеличены у животных после перерезки пуповины (рис. 55.2 (c)) по сравнению с возрастом — связанные контроли, достигающие значений, близких к тем, которые наблюдаются у контрольных животных на P1–3 (рис. 55.2 (a)). После ежедневных инъекций агониста 5-HT 2 R DOI иммуномечение GABA A R на MN P7 после SCT при рождении упало до значений, указанных в контроле-P7, в то время как маркировка GlyR оставалась стабильной.ТСК, проведенная на P5, когда уровни GABA A R уже являются низкими в поясничном отделе спинного мозга, значительно усилила экспрессию GABA A R через 1 неделю (то есть такая же задержка между пересечением спинного мозга и анализом, как и в экспериментах с рассечением при рождении) с небольшим влиянием на GlyR. Эти эксперименты демонстрируют, что пластичность развития GlyR не зависит от супраспинальных влияний, тогда как на пластичность GABA A R заметно влияют нисходящие пути, в частности серотонинергические проекции, поскольку лечение агонистом 5-HT 2 R имитировало нормальное развитие (Садлауд и др., 2010).
Рефлекс растяжения и миф о взаимном «торможении» — НЕЙРОПЕДИЧЕСКАЯ РЕАБИЛИТАЦИЯ И ДВИЖЕНИЕ КОНСАЛТИНГОВАЯ КОМПАНИЯ
Понимание того, что происходит в неврологически неблагополучных ситуациях, может улучшить наше практическое понимание ортопедических случаев.
В неврологически измененной ситуации, например, при инсульте или травме головного мозга, нервные пути нисходящей модуляции повреждаются, и по какой-то причине это делает волокна веретена чрезвычайно чувствительными практически к любым раздражителям (включая эмоции или ночь минимального сна).Клинически мы наблюдаем серьезную аномальную тональную синергию в конечностях, которая в некоторых случаях может сделать произвольное функциональное использование конечностей практически невозможным.
Если вы работали с пациентами, у которых истинный неврологический тонус вторичен по причине травмы головного мозга, вы знаете, что ничего не добьетесь, если попытаетесь «растянуть» и бороться с этим тонусом, имея в виду, что вы собираетесь механически «удлинить его». , »Иногда это действительно ухудшает тон.
В неврологических условиях мы можем достичь глубокого функционального использования конечностей, если переключим фокус на оптимизацию контроля над проксимальными сегментами туловища i.лопатка и таз, периодически выполняя изометрические периферические упражнения.
То же самое применимо в ортопедической ситуации, когда ограниченное ПЗУ связано с повышением неврологического тонуса. Например, вместо того, чтобы пытаться растянуть подколенные сухожилия у кого-то с болью в пояснице с намерением пассивно удлинить ткань, мы видим лучшие результаты при выполнении прогрессивных угловых изометрических упражнений на подколенных сухожилиях с периодическими упражнениями на стержень против растяжения для оптимизации проксимальной стабильности. для дистальной подвижности.
С неврологической точки зрения «тугие подколенные сухожилия» могут быть одним из единственных способов для ЦНС решить «проблему степеней свободы» при плохом контроле пояснично-тазового туловища. «Проблема степеней свободы» родилась из модели динамических систем управления моторикой, представленной Николаем Бернштейном четвертой и широко обсуждаемой в этой статье: Теория динамических систем развития моторики: Полное руководство для специалистов по спортивной реабилитации.
Клинически д-р Кабат и Мэгги Нотт использовали нейрофизиологические принципы волокон веретена, основанные доктором М.Шеррингтоном в 1930-х годах: вызвать произвольные сокращения мышц или снизить ненормальный тонус с помощью методов, помогающих «перенастроить» уровни чувствительности веретена у тех, кто имел неврологические нарушения. Понимание рефлекса растяжения на этом уровне может помочь переосмыслить наши клинические взгляды на улучшение мобильности и / или контроля над моторикой у наших клиентов.
В клинических, неврологических или ортопедических условиях, когда мы видим такие же улучшения сеанса с тестом перед пост-постом движений, это просто проявление перекалибровки рефлекса растяжения ЦНС.Независимо от того, используем ли мы изометрические методы, стратегии проксимальной стабилизации или проблемы с осанкой, мы можем напрямую влиять на модуляцию рефлекса растяжения в ЦНС, чтобы оптимизировать подвижность и моторный контроль.
ссылки
1. Мур М.А., Хаттон Р.С. Электромиографическое исследование техники растяжения мышц. Med Sci Sports Exercise 1980; 12: 322-9.
2. Магнуссон С.П., Симонсен Э.Б., Аагард П., Дайре-Поулсен П., МакХью М.П., Кьяер М. Механические и физиологические реакции на растяжение с предварительным изометрическим сокращением и без него в скелетных мышцах человека.Arch Phys Med Rehabil 1996; 77: 373-8.
3. Маркос П.Д. Ипсилатеральные и контралатеральные эффекты проприоцептивных нейромышечных методов облегчения движения бедра и электромиографической активности. Phys Ther 1979; 59: 1366-73.
4. Остерниг Л. Р., Робертсон Р., Троксель Р., Хансен П. Активация мышц во время проприоцептивных методов растяжения нервно-мышечной системы (PNF). Am J Phys Med 1987; 66: 298-307.
5. Halbertsma JPK, Goeken LNH. Упражнения на растяжку: влияние на пассивную растяжимость и жесткость коротких подколенных сухожилий здоровых людей.Arch Phys Med Rehabil 1994; 75: 976-81.
Работа сэра Чарльза Шеррингтона «Интегративное действие нервной системы: признание к столетию» | Мозг
Аннотация
В 1906 году сэр Чарльз Шеррингтон опубликовал «Интегративное действие нервной системы», представлявший собой сборник из десяти лекций, прочитанных двумя годами ранее в Йельском университете в США. В этой монографии Шеррингтон резюмировал два десятилетия кропотливых экспериментальных наблюдений и свою проницательную интерпретацию их.Он уладил тогдашние споры между идеями «ретикулярной теории» и «нейронной доктрины» о фундаментальной природе нервной системы млекопитающих в пользу последних и навсегда изменил то, как последующие поколения рассматривали организацию центральная нервная система. В большом произведении Шеррингтона содержатся основные концепции и даже терминология, которые теперь стали второй натурой для каждого изучающего этот предмет. В этой краткой статье рассматривается исторический контекст, в котором была написана книга, резюмируется ее содержание и рассматривается ее влияние на неврологию и нейробиологию.
Введение
Первое десятилетие 20-го века ознаменовалось двумя важными событиями для науки. 1905 год был «чудесным годом» Альберта Эйнштейна, в течение которого были опубликованы три его самые знаменитые работы по теоретической физике. Следующий 1906 год был не менее важен для неврологии и нейробиологии. Нобелевская премия по физиологии и медицине была присуждена двум великим нейроанатомам, итальянцу Камилло Гольджи и испанцу Сантьяго Рамон-и-Кахаль.Их противоборствующие Нобелевские лекции олицетворяли нарастающий триумф «нейронной доктрины», поддерживаемой Кахалем, над «ретикулярной теорией», которую горячо поддерживал Гольджи (; стр. 29–30). Анатомические наблюдения и идеи Кахаля, обобщенные в его монументальном Histologie du système nerveux de l’homme et des vertébrés (Ramon y Cajal, 1909, 1911, 1995), уже вдохновили молодого британского физиолога Чарльза Скотта Шеррингтона взглянуть на Центральная нервная система млекопитающих совершенно по-новому.В результате в 1906 году Шеррингтон опубликовал интегративного действия нервной системы (Sherrington, 1906), в котором обобщены почти два десятилетия его интенсивных исследований и размышлений. Без преувеличения можно сказать, что книга Шеррингтона изменила последующий курс нейрофизиологии. Эта записка предназначена для празднования столетия этого знаменательного события.
Лекции Силлимана
Монография Шеррингтона 1906 года, опубликованная одновременно в Лондоне, Нью-Хейвене и Нью-Йорке, была основана на серии из 10 специальных лекций, прочитанных в 1904 году в Йельском колледже в Нью-Хейвене, Коннектикут, под эгидой Фонда Силлимана.По завещанию Силлимана, Йельский колледж был
… запрошен и направлен на организацию ежегодного курса лекций, предназначенных для иллюстрации присутствия и провидения, мудрости и доброты Бога, проявленных в естественном и моральном мире. (Неподписанное предисловие к изданию 1906 г.)
Эта директива, рассмотренная более чем через столетие, кажется странной, но она идеально подошла профессору Шеррингтону. Он был тогда и позже в жизни не только опытным экспериментатором и наблюдателем, но также поэтом и философом, а также настоящим гуманистом в лучшем смысле этого слова.
Шеррингтон был вторым лектором Силлимана, первым из которых был профессор Дж. Дж. Томсон из Кембриджского университета, который говорил на тему «Электричество и материя». После двухлетней задержки (за которую Шеррингтон извиняется в своем кратком предисловии) он подготовил лекции к публикации в виде монографии. Интегративное действие впоследствии переиздавалось четыре раза между 1906 и 1920 годами и переиздавалось в 1947 году издательством Кембриджа и Йельского университета (Sherrington, 1947) с новым предисловием Шеррингтона.В этом предисловии Шеррингтон написал обширное философское эссе «… чтобы разобраться с некоторыми неясностями, которые со временем возникли» (стр. Ix в издании 1947 года). Вот как Шеррингтон, которому тогда было 90 лет, увидел свою монографию через 40 лет после ее первого появления:
Переизданный здесь том касается преимущественно того типа моторного поведения, который называется «рефлексом»; может создаться впечатление, что в рефлекторном поведении он видит самый важный и далеко идущий из всех типов «нервного» поведения.На самом деле это не так. Но рефлекторное действие дает определенные преимущества для физиологического описания. Его можно изучать без осложнений из-за того типа «нервной» активности, который называется автохтонным (или «спонтанным») и порождает внутренне возникающие ритмические движения, например дыхание и т. д. Но по сравнению с обширной областью поведения в целом, чистое рефлекторное действие само по себе не может покрывать такую обширную область, как инстинкты, вызванные «побуждениями» и «побуждениями». Но механизм их еще недостаточно изучен для лабораторного лечения.Чистый апсихический рефлекс играет меньшую роль. Изученное в этой замкнутой группе животных, позвоночных, поведение, кажется, становится все менее и менее рефлекторным по мере того, как особь животного становится все более и более полностью индивидуализированной. «Спинальный» человек более искалечен, чем «спинномозговая» лягушка. (стр. ix – x в перепечатке 1947 года.)
Наконец, издание 1947 года было снова полностью переиздано в 1961 году в виде тома в бумажном переплете, выпущенного издательством Йельского университета. Эта замечательная история публикаций свидетельствует не только о знаковой классике физиологии, но и о неизменной актуальности идей Шеррингтона о том, как работает центральная нервная система.
Источники
Несмотря на то, что, как говорит нам Лидделл, «… Шеррингтон неоднократно заявлял с большой серьезностью, что надеется, что никто не напишет его биографию» (Liddell, 1960, предисловие), его коллеги и бывшие студенты предоставили нам завидное количество статей и книги, которые передают факты его жизни и комментарии к его подходу к науке и миру в целом (Adrian, 1957; Cohen, 1958; Denny-Brown, 1957; Eccles, 1957, 1982; Eccles and Gibson, 1979; Granit, 1966 г .;).Существуют также более свежие оценки научного вклада Шеррингтона в контексте современных идей в нейрофизиологии (Swazey, 1969; Stuart et al., ., 2001; Breathnach, 2004; Stuart, 2005). После его смерти в 1952 году в возрасте 94 лет появились многочисленные некрологи, снова написанные бывшими студентами и выдающимися коллегами, которые содержат важную информацию о жизни и мысли Шеррингтона (см. Избранные ссылки в Eccles and Gibson, 1979). Обширный некролог Лидделла (Liddell, 1952) особенно красноречив и информативен.Эти сочинения выражают глубокую и искреннюю привязанность к Шеррингтону как к человеку, а также как к научному наставнику. Особо следует отметить том, озаглавленный The Discovery of Reflexes , автор E.G.T. Liddell (), в которой представлен замечательный обзор научного наследия, сформировавшего мышление Шеррингтона в то время, когда он читал свои лекции по Силлиману.
Наконец, Дерек Денни-Браун, студент Шеррингтона, ставший выдающимся клиническим неврологом, составил Избранные труды сэра Чарльза Шеррингтона с комментариями, впервые опубликованные в 1939 году Хэмишем Гамильтоном для гарантов мозга и перепечатанные издательством Oxford University Press в 1979 (Денни-Браун, 1979).Эта бесценная коллекция включает в себя научные труды Шеррингтона, многие из которых трудно получить в оригинале, и заслуживает места на полке каждого нейробиолога.
Немного фона
Чарльз Скотт Шеррингтон родился 27 ноября 1857 года в английской семье среднего достатка. Он получил медицинское образование в Кембридже и в больнице Святого Томаса в Лондоне. Во время учебы в Кембридже Шеррингтон занял первое место в обеих частях экзаменов по естественным наукам и справился, несмотря на свой миниатюрный рост (5 футов 6 дюймов; см. Granit, 1966; стр.3), и в то же время быть серьезным конкурентом своего колледжа как по регби, так и по гребле. Его первые две статьи были опубликованы в 1884 году вместе с его кембриджским наставником Дж. Н. Лэнгли по гистологическим исследованиям мозга собак, подготовленным профессором Гольцем из Страсбурга (Denny-Brown, 1979). Позже Шеррингтон провел с Гольцем более полугода, выполняя анатомические работы и впитывая идеи Гольца о локализации функции или ее отсутствии в коре головного мозга.
Получив квалификацию врача в 1885 году, Шеррингтон заинтересовался исследованиями инфекционных заболеваний, таких как дифтерия и холера, и их улучшения с помощью недавно разработанных вакцин.В том же году он присоединился к своим друзьям К. С. Рою и Дж. Грэму Брауну в летней экскурсии в Испанию, чтобы исследовать предполагаемую вакцину от разразившейся там азиатской холеры. Хотя сообщения различаются (например, Liddell, 1952; стр. 245), Эклс и Гибсон (1979; стр. 4) заявляют, что Шеррингтон не встречался с Рамоном-и-Кахалем во время той поездки, отмечая, что его единственная прямая встреча с великим испанским нейроанатомом имела место позже, во время визита Кахала в Англию в 1894 году. Следующим летом Шеррингтон отправился в аналогичную экспедицию в Италию, результатом которой стал длительный визит в Берлин с выдающимся патологом Рудольфом Вирховым.Вирхов отправил его в лабораторию Роберта Коха, одного из ведущих бактериологов того времени, где он провел целый год (Гранит, 1966; с. 15).
После возвращения в Англию в 1887 году Шеррингтон был назначен лектором по физиологии в больнице Святого Томаса и был избран в стипендию в колледже Гонвилля и Кая в Кембридже. Он снова обратился к невропатологическим исследованиям человеческого мозга и опубликовал ряд отчетов на эту тему в течение следующих 5 лет. Именно в этот период интересы Шеррингтона переместились с коры головного мозга на спинной мозг с использованием экспериментальной физиологии, а также нейроанатомических методов.В этом на него большое влияние оказал В. Х. Гаскелл, преподававший на кафедре физиологии сэра Майкла Фостера в Кембридже. Позже, в 1918 году, Шеррингтон написал своему другу Генри Хэду, что «моя собственная работа началась случайно не с того конца — корковые пирамидальные дегенерации и т. Д.… [Он] не мог поговорить с ним [Гаскеллом]». долго, не осознавая, что пуповина представляет собой лучшую точку атаки с физиологической точки зрения »(цитируется по Liddell, 1952; стр. 244).
В 1891 году Шеррингтон был назначен суперинтендантом Института Брауна Лондонского университета, ветеринарной больницы, которая предоставила ему большой материал для экспериментальной работы.В это время он был избран членом Королевского общества и побудил эту организацию пригласить профессора Кахала для чтения крооновской лекции в 1894 году. Приключения Кахаля в Англии незабываемо описаны в биографии Шеррингтона Экклза и Гибсона (1979; стр. 10). –12).
Именно в этот период в Институте Брауна окончательно проявился страстный интерес Шеррингтона к физиологии нервной системы. Между 1889 и 1895 годами он опубликовал серию статей, число которых постоянно увеличивалось, и в них стали входить конкретные работы по рефлексам.Особое значение имели две статьи о коленном рефлексе (Sherrington, 1893, a , b ), в которых он представил ключевые доказательства того, что это хорошо известное клиническое явление действительно было рефлексом (тогда в некоторых спорах), который зависел от афферентов. возникает в сокращающихся мышцах-разгибателях колена, и эта активация афферентов от мышц-сгибателей коленного сустава-антагониста уменьшала или отменяла толчок разгибателя. Он воспользовался наличием в институте больших и малых приматов, а также других видов, чтобы детально проработать сегментарные паттерны афферентной и эфферентной иннервации определенных мышц, а также межжелезные вариации, которые имели решающее значение для интерпретации более поздних рефлекторных исследований (Sherrington, 1892, 1894).
Шеррингтон был приглашен в 1895 году на должность профессора физиологии Университетского колледжа Ливерпуля. Здесь он должен был оставаться до 1914 года, когда он уехал, чтобы занять кафедру физиологии в Оксфорде. Хотя последний период более известен, особенно благодаря комментариям выдающихся студентов, которые работали с ним в эпоху Оксфорда (Denny-Brown, 1957; Eccles, 1957, 1982; Eccles and Gibson, 1979; Granit, 1966), друга Шеррингтона и его коллега лауреат Нобелевской премии Э.Д. Адриан прокомментировал, что «… когда он занимал кафедру физиологии в Ливерпуле, он был на пике своих возможностей как исследователь» (Adrian, 1957;212). Сын Шеррингтона Карр написал, что оба его родителя нашли жизнь в Ливерпуле гораздо более благоприятной, чем жизнь в Лондоне: «… годы в Ливерпуле, я считаю, были самыми счастливыми в его жизни, и моя мать тоже…» (см. Приложение 17 в Eccles and Gibson, 1979). В той же статье показано, насколько занят Шеррингтон был как снаружи, так и внутри лаборатории в период вынашивания ребенка Интегративное действие (см. Также стр. 127–143).
Что сказал Шеррингтон в 1906 году?
Обзор
Интегративное действие нервной системы состоит из 10 глав, каждая из которых охватывает одну из лекций 1904 года в Йельском университете.Книга была посвящена Дэвиду Ферьеру, одному из научных героев Шеррингтона, «… в знак признания его многочисленных заслуг в экспериментальной физиологии центральной нервной системы». Шеррингтон в своем предисловии объяснил двухлетнюю задержку «давлением». разнообразной работы…. Пользуясь случаем, хочу выразить сожаление по поводу задержки ». Из-за формата лекции в последующих главах есть определенное количество повторений. Кроме того, уникальный стиль письма Шеррингтона, несомненно, под влиянием его образования в классической литературе и его навыков поэта, иногда бывает трудным.Лорд Адриан отметил в честь столетнего юбилея Шеррингтона: « Интегративное действие нервной системы … это не легкое чтение, но оно есть и останется одним из основных классических произведений физиологической литературы» (Adrian, 1957). ; с. 214).
Ниже представлена попытка обобщить содержание книги с точки зрения того времени, когда она была опубликована. Каждой из 10 лекций предшествует «аргумент», в котором Шеррингтон кратко излагает основные моменты, на которые следует обратить внимание.Текст включает многочисленные цитаты из предыдущих работ других авторов, а также его самого. В библиографии 314 ссылок, от Декарта в 17 веке до начала 20 века, включая цитаты на французском, немецком, итальянском и испанском языках, во всех из которых Шеррингтон был грамотным. Приведенные ниже ссылки на страницы относятся к переизданию 1961 года, потому что это, вероятно, наиболее доступная для нынешних читателей версия.
Лекции
Первые три лекции посвящены «Координации простого рефлекса».Шеррингтон смело формулирует общий смысл всей монографии в знаменитом первом предложении лекции I: «Нигде в физиологии клеточная теория не обнаруживает своего присутствия в самой структуре аргументации так часто, как в настоящее время при изучении нервной системы. реакции ». С самого начала своей научной карьеры Шеррингтон был убежден, что нейроны — это отдельные сущности, а не части непрерывной сети напрямую связанных каналов, по которым сигналы проходят беспрепятственно, как это предусмотрено« ретикулярной теорией ».Он представляет точку зрения, согласно которой центральная нервная система «… работает через живые линии неподвижных клеток, по которым она посылает волны физико-химических нарушений… относительно высокой скорости…» (стр. 3 перепечатки 1967 года). Он определяет рефлекторную дугу как «… целую цепочку структур — рецептор, проводник и эффектор…» и продолжает утверждать, что «единичной реакцией в нервной интеграции является рефлекс» (стр. 7). Шеррингтон сразу же квалифицировал понятие простого рефлекса как «… вероятно, чисто абстрактное понятие, потому что все части нервной системы связаны друг с другом, и ни одна из ее частей, вероятно, никогда не способна реагировать с воздействием и воздействием на различные другие части, и это система, определенно никогда не находящаяся в состоянии покоя.И, наконец, он добавляет: «Главный секрет нервной координации, очевидно, заключается в сложении рефлексов» (стр. 8). Таким образом, на нескольких страницах Шеррингтон представляет свою широкую концепцию центральной нервной системы, которая составляет основу остальной части монографии.
Остальная часть первой лекции посвящена таким важным концепциям, как специфичность сенсорных рецепторов, центральная латентность проводимости в рефлекторных дугах, которая медленнее, чем сама нервная проводимость, однонаправленная проводимость в отличие от двунаправленной проводимости в нервных волокнах, постразряд, временное суммирование при повторяющихся раздражителях, рефлекторная усталость, порог рефлекса и его вариабельность, а также зависимость рефлексов от нормального метаболизма и эффекта таких лекарств, как анестетики.Он использует все эти особенности, чтобы подчеркнуть, что должна быть «… связь между нейроном и нейроном…» (стр. 17), то есть «синапс», который отвечает за однонаправленное проведение в рефлекторных дугах. Он отмечает, что ввел этот термин ранее в издание 1897 года Учебника физиологии сэра Майкла Фостера . Шеррингтон иллюстрирует свои экспериментальные доказательства всех «аргументов» на протяжении первых трех лекций подробными иллюстрациями своих миографических записей. Текст украшен ссылками на существующую литературу с примерами от медуз до собак и кошек.
Лекция II продолжает изложение Шеррингтоном контрастов между рефлекторным действием и тем, что можно ожидать от проведения через ретикулярную сеть, снова подчеркивая свойства необратимости, суммирования подсознательных стимулов, облегчения последовательных стимулов и длительных действий, являющихся результатом кратковременных стимулы. Используя в качестве примера рефлекс царапания, он отмечает важность «рефрактерной фазы», во время которой последующие стимулы не вызывают рефлекса.В качестве яркого свидетельства своих представлений о центральной организации рефлекторных путей он приводит предлагаемую принципиальную схему рефлекса растяжения (рис. 13B), которая включает три последовательных нейрона: афферентный «рецептивный нейрон» и длинный нисходящий «проприоспинальный нейрон». и двигательный нейрон «конечного общего пути». Он предположил, что в этой цепи есть два центральных синапса, и поэтому описывает ее как «дисинаптическую дугу» (стр. 53). Такое конкретное предложение было бы буквально немыслимо в ретикулярной нервной системе.Поскольку ни афференты, ни мотонейроны не проявляют длительного рефрактерного периода в рефлексе царапания, Шеррингтон заключает, что «… место рефрактерной фазы рефлекса царапания лежит»… в самом центральном нервном органе, где-то между двигательным нейроном и нервным нервом. мышца и рецептивный нейрон из кожи »(стр. 65). Современный читатель простит предположение Шеррингтона о том, что последовательные рефрактерные фазы объясняют циклические движения конечностей при царапании и « вероятно » (стр.65) шагающий. Как отмечалось позже, его взгляд столетней давности не включал центральные схемы, которые могут создавать такие повторяющиеся паттерны движений.
В заключительном разделе лекции III Шеррингтон вводит то, что он, возможно, считал своим величайшим открытием, — взаимное торможение (Гранит, 1966; с. 50). Он определил это как рефлекс «одновременного двойного знака» с возбуждением одной или нескольких мышц-агонистов и одновременным ингибированием антагонистов, действующих на один и тот же сустав. Он смог показать, что локус этого торможения находится в спинном мозге, а не на периферии, как это было обнаружено другими у некоторых беспозвоночных.Шеррингтон описывает реципрокное торможение, когда оно накладывается на преувеличенный тонус разгибателей, обнаруживаемый в «децеребрационной ригидности» (еще один неологизм Шеррингтона, который все еще используется сегодня), а также в фазическом коленном рефлексе и других рефлексах задних конечностей. Не менее важно то, что он продемонстрировал, что задействованные афференты перемещаются по нерву, который иннервирует отвечающую мышцу. Такие афференты, по его терминологии, являются «проприоцептивными», потому что они активируются собственными движениями организма, в отличие от «экстероцептивных» афферентов, передающих информацию из окружающей среды.Шеррингтон тщательно описывает предостережение о том, что при тестировании на наличие реципрокного торможения необходимо учитывать механическое действие пар мышц-агонистов / антагонистов, чтобы избежать ошибок при взгляде на мышцы, пересекающие два сустава (те, которые сейчас известны как бифункциональные или многофункциональные мышцы). . Лекция заканчивается тестами на судорожный препарат стрихнин и действие столбнячного токсина. Он комментирует, что его наблюдения «… склоняют меня к выводу, что действие алкалоида заключается в преобразовании в спинном мозге процесса торможения — каким бы он ни был по существу — в процесс возбуждения — чем бы он ни был по существу» (стр. .112).
В лекции IV Шеррингтон начинает свое рассмотрение взаимодействия рефлексов, которое, как он отмечает, является «… главной проблемой нервной координации», и вводит то, что он называет «… принципом общего пути» (стр. 117). . Он различает «частные» пути от индивидуальных сенсорных рецепторов и нейронов в спинном мозге, которые в разной степени представляют «общественные» пути, то есть нейроны, которые получают входные данные, сходящиеся от множества афферентов. Что касается моторных рефлексов, мотонейроны, активирующие мышцы, обязательно представляют собой « последний общий путь », но Шеррингтон старается отметить, что « внутренние нейроны » (теперь чаще называемые интернейронами) также представляют собой общие пути, которые получают конвергентные сигналы не только от афферентов, но также из других наборов интернейронов.На этом фоне он исследует экспериментальные результаты рефлексов, которые «… действуют гармонично вместе…», называемых «союзными рефлексами», в отличие от «антагонистических рефлексов», которые исключают или прерывают друг друга. Поскольку все они действуют через один и тот же общий путь final , мотонейроны, он утверждает, что центральные механизмы должны включать взаимодействия между общими межнейронными путями, которые получают различные комбинации афферентных входов. Он проводит резкий контраст между расхождением афферентных входов, когда они входят в спинной мозг, и конвергенцией этих входов на нейроны по множеству общих путей, которые в конечном итоге питаются мотонейронами, которые являются последним общим путем.Шеррингтон представляет данные, чтобы показать, что родственные рефлексы одного и того же «типа» (например, рефлекс царапания из разных частей рецептивного поля) демонстрируют подкрепление, или то, что он называет «немедленной индукцией позвоночника», которая увеличивается по мере уменьшения расстояния между стимулируемыми точками. Хотя он отмечает, что «… интимную природу механизма… трудно предположить…» (стр. 143), Шеррингтон использует принципиальные схемы (рис. 38, повторение рис. 13 в лекции II и 44), чтобы проиллюстрировать, как межнейронный конвергенция может быть основой для положительных и отрицательных рефлекторных взаимодействий.Именно размышления Шеррингтона о центральных нейронных путях трансформировали феноменологию в науку нейрофизиологию, которой мы следуем по сей день.
Лекция V продолжает обсуждение Шеррингтоном взаимодействий между рефлексами как отражений внутриспинальных цепей, с идеей синаптической силы как ключа к наблюдаемым пороговым различиям в рефлекторных взаимодействиях (его рис. 46). Он формулирует пять «правил, участвующих в распространении импульсов в спинальных рефлексах», которые в основном касаются сегментарной локализации афферентных и эфферентных конечностей дуги.Он также развенчивает «четыре закона» Пфлюгера, относящиеся к одному и тому же предмету (см. Стр. 84–87), заявляя, что «эти так называемые« законы »рефлекторного излучения были настолько общепринятыми, что приобрели доктринальную значимость. чего они вряд ли заслуживают »(стр. 165). Это один из немногих случаев, когда Шеррингтон, обычно сдержанный в подобных вопросах, вступает в откровенно полемический спор. Он обсуждает свою концепцию «рефлекторных фигур», в которой поза животных с надспинальными перерезками на бульбарном или спинномозговом уровнях объясняется одновременным сочетанием «гармоничных» длинных и коротких дуговых рефлексов.Шеррингтон также опровергает широко распространенное (в то время) мнение о том, что такие гармоничные комбинации можно объяснить комбинациями отдельных спинных корешков (то есть сегментов).
В лекции VI Шеррингтон переходит от идеи одновременных рефлексов к координации через временные последовательности рефлексов, приближаясь к своей цели — объяснению роли рефлексов в моторном поведении у ведущих себя млекопитающих. Он подчеркивает, что комбинации последовательных рефлексов могут включать как «союзнические», так и «антагонистические» рефлексы, которые «… связаны друг с другом более чем просто внешним… стимулом.В такой последовательности порог каждого последующего рефлекса понижается возбуждением, непосредственно предшествующим его собственному . ‘(Стр. 185; курсив в оригинале), что приводит к плавным переходам, которые производят скоординированное движение. В середине лекции (стр. 196–200) Шеррингтон внезапно обращается к увлекательному анализу современных представлений о механизме центрального торможения, которые имеют удивительно современный тон. Очевидно, такие внезапные отступления были не редкостью в лекциях Шеррингтона (Гранит, 1966; с.19; см. также Eccles and Gibson, 1979; п. 46).
Особый интерес для современных читателей представляет краткое обсуждение Шеррингтоном «шагового рефлекса», под которым он подразумевает последовательное разгибание и сгибание конечности в позвоночнике собаки, когда она подвешена над землей. Он упоминает это как пример адаптации рефлексов к определенному действию, в данном случае к локомоции. Его обсуждение предполагает, но не указывает четко, что он рассматривал это действие как зависимое от проприорецепторов конечностей, поскольку оно может происходить без очевидного внешнего стимула.Он также отмечает, что «у нас пока нет удовлетворительного объяснения этого». Другие (Stuart et al ., 2001) недавно отметили, что кажется странным, что Шеррингтон в значительной степени игнорировал проблему автономного шагания по позвоночнику в 1906 году, а также позже. , хотя эта проблема была исследована Томасом Грэмом Брауном в новаторской работе, начатой в департаменте Шеррингтона в Ливерпуле (Graham Brown, 1916). Ответ можно найти в философском предисловии Шеррингтона к переизданию 1947 года книги The Integrative Action (стр.x, процитировано выше), в котором он сознательно исключил такие «автохтонные» действия из рассмотрения.
Внутренне генерируемые ритмические движения действительно представляют собой действия, которые являются результатом сложных нейронных цепей, что ясно демонстрирует текущая работа над тем, что сейчас называется «генераторами центральных паттернов». Шеррингтон, очевидно, считал, что рефлекс царапания, которому уделяется много внимания в Интегративное действие , принципиально отличается (хотя явно представляет собой сложное ритмическое действие), потому что он запускается внешним стимулом, как и любой другой рефлекс.В настоящее время считается, что ритмическое царапанье является запускаемой реакцией, в которой задействован центральный генератор паттернов, тесно связанный с этим движением. У млекопитающих эти центральные цепи все еще неуловимы, несмотря на современные методы электрической и оптической записи с одного или нескольких нейронов. Они были совершенно недоступны, учитывая относительно примитивные экспериментальные подходы, доступные во времена Шеррингтона. Неудивительно, что Шеррингтон даже в 1947 году предпочел придерживаться своей точки зрения, согласно которой «… последовательность двигательных действий, следовательно, является результатом последовательности последовательных внешних ситуаций» (стр.xi).
В своей замечательной лекции VII Шеррингтон рассматривает три разных вопроса. Во-первых, он решает запутанную телеологическую проблему «почему» рефлексов. Отвергая более ранние представления о «психических силах спинного мозга», он предполагает, что дарвиновская «адаптация под естественным отбором» обеспечивает объяснение существования «целенаправленных нейронных механизмов» (стр. 236 и 237). Он сочетает убедительные философские аргументы с множеством примеров, чтобы проиллюстрировать свой случай. Затем он переходит к рассмотрению природы спинномозгового шока, который следует за перерезкой позвоночника, отмечая относительное отсутствие изменений над поражением в отличие от драматических изменений под ним.Он отмечает, что это явление гораздо более глубокое и продолжительное у приматов, чем у «низших» видов, и что оно влияет на ноцицептивные рефлексы гораздо меньше, чем другие, такие как рефлекс царапания. После краткого обсуждения тонких различий, зависящих от локуса стимула, называемого «локальным знаком», Шеррингтон начинает расширенное эссе о нейронной основе эмоции. Он определяет «псевдоаффективные рефлексы» как реакцию децеребрирующих животных на стимулы, которые, как ожидается, вызовут боль, если животное не повреждено.Внешние признаки подобны тем, которые наблюдаются у интактных животных, но «… они никогда не сводятся к эффективному действию нападения или побега» (стр. 253). Используя повреждения позвоночника в таких препаратах, Шеррингтон приходит к выводу, что ноцицептивные пути восходят к мозгу только через латеральные столбики, больше через те, которые контралатеральны стимулу, чем через ипсилатеральные. Он также заключает, используя данные других исследователей (в частности, Гольца), что центры, участвующие в ноцицепции, расположены ниже в нейраксисе, чем центры, предназначенные для восприятия удовольствия.Это приводит его к обширному обсуждению трех современных теорий эмоционального восприятия: (1) внешний стимул вызывает эмоциональное восприятие в мозгу, которое, следовательно, производит висцеральное (вегетативное) сопровождение; (2) стимул порождает восприятие и висцеральные реакции одновременно или (3) стимул вызывает висцеральные реакции, которые, в свою очередь, воспринимаются мозгом как эмоциональное восприятие, как утверждали Уильям Джеймс и, несколько иначе, Ланге и Серджи. .Шеррингтон использует свою модель псевдоаффективного рефлекса, чтобы изучить вопрос у животных с перерезками позвоночника и блуждающего нерва. Результаты экспериментов описаны очень подробно, что привело Шеррингтона к выводу, что «… внутреннее выражение эмоции на вторично по отношению к мозговому действию, происходящему с психическим состоянием» (стр. 266; курсив в оригинале). В одной лекции Шеррингтон делает смелый прыжок от подробного описания внутренней работы рефлексов в спинном мозге к гораздо более философскому и, в некоторой степени, спекулятивному эссе о «высшей нервной функции» мозга.
В лекции VIII Шеррингтон завершает свое восхождение к нейраксису подробным обсуждением областей «моторной коры», которые, как он и другие показали, производят определенные движения при электрической активации в определенных и воспроизводимых корковых локусах. Он резюмирует свою работу в Институте Брауна с множеством человекообразных обезьян, подчеркивая, что такие локусы могут быть точно нанесены на карту, даже если они ненадежно связаны с корковыми бороздами и извилинами (за исключением центральной борозды) у отдельных животных.Большая часть этой лекции посвящена вопросу реципрокного торможения реакций, вызываемых корковой стимуляцией. Шеррингтон подчеркивает, что паттерны реципрокного торможения между «истинными» парами антагонистов во многом одинаковы, независимо от того, вызвано ли оно корковой стимуляцией или периферическими входами в спинной мозг. Несмотря на то, что стимуляция коры обычно способствует возбуждению групп сгибателей, происходит соответствующее ингибирование разгибателей-антагонистов. Шеррингтон использует сходные эффекты стрихнина и столбнячного токсина на сегментарные (так называемые «местные») рефлексы и ответы на корковую стимуляцию, чтобы постулировать, что реципрокное торможение, обнаруженное в обоих, осуществляется сегментарными механизмами.В последней части лекции дается подробное описание децеребральной ригидности, которую он использует для постулирования «двух разделимых систем моторной иннервации…» (стр. 312), обе из которых требуют целых афферентов: «тоническая» система, активирует антигравитационные мышцы по всему телу, а другая — «фазовую» систему, которая производит множественные «локальные» рефлексы, обсуждавшиеся в предыдущих лекциях. Он размышляет о дихотомии между корой и мозжечком, так что «… мозжечок является центром непрерывных движений, а мозг — изменением движений» (стр.303).
Лекция IX, озаглавленная «Психологическое положение и доминирование мозга», представляет собой эссе tour-de-force об общих принципах организации центральной нервной системы в животном мире, от простейших до человека. В нем Шеррингтон отходит от деталей, которые занимают большую часть предыдущего материала, чтобы представить грандиозную панораму нервных систем через филогенетическую последовательность. Он объединяет идеи, развитые в предыдущих лекциях, резюмированные с исключительной ясностью.Вначале Шеррингтон отмечает, что «… своим разветвлением двигательный нейрон захватывает многие мышечные волокна» (стр. 309), что является основной идеей, лежащей в основе его более позднего введения термина «двигательная единица» (Liddell and Sherrington, 1925). ). Развивая уже обсуждавшиеся идеи об экстероцептивных и проприоцептивных афферентах, а также об организации спинных сегментов и основных двигательных рефлексах, Шеррингтон представляет критическую важность лабиринтов как датчиков положения в пространстве, действующих совместно с проприоцептивной системой, управляемой ею, и являющейся ее частью. мозжечок.Однако его цель — разъяснить роль полушарий головного мозга, называемых «мозгом». Шеррингтон рассматривает дистанционные рецепторы (включая, помимо прочего, зрительную, слуховую и обонятельную системы) как необходимые для успеха организма в выживании в окружающей его среде. Это мозг, который объединяет их информацию, чтобы направлять действия «последующих сегментов». Он говорит: «Мозг всегда является частью нервной системы, которая построена на« органах-рецепторах расстояния »и развивается на их основе» (стр.325). Здесь так много всего, что невозможно подвести итоги. Если в «Интегративное действие » больше ничего не читается, то это должна быть эта лекция. Любой, у кого есть хотя бы мимолетный интерес к мозгу, найдет здесь чудесное переплетение фактов и убедительного анализа, выраженное красивым языком.
Заключительная лекция, озаглавленная «Чувственное слияние», на первый взгляд может показаться чем-то вроде разочарования после вышеизложенного, но она представляет собой попытку Шеррингтона выйти за пределы моторной системы в сферу психических событий, т.е.е. ощущение и восприятие. Вначале он говорит:
. Но мы можем согласиться с тем, что если такие ощущения и чувства или что-то подобное им действительно сопровождает реакции, которые мы изучали, то нервный механизм, к работе которого они присоединяются, не ограничен нервными дугами. мы до сих пор прослеживали, но в областях нервного аппарата, которые, хотя и связаны с этими дугами, лежат за ними, в полушариях головного мозга (стр. 353–354).
Будучи экспериментатором, он начинает с подробного описания своей работы по восприятию контраста в мерцающих зрительных стимулах, предъявляемых одновременно к обоим глазам.В отличие от своей ясности в предыдущей лекции, Шеррингтон описывает свой аппарат и результаты длинного списка вариантов протокола на эллиптическом языке, что затрудняет чтение. Однако затем он поясняет свою цель, которая состоит в том, чтобы показать, что наблюдаемые эффекты обусловлены не периферическими взаимодействиями, а скорее явлениями, возникающими в результате:
… только после того, как ощущения, инициированные из правой и левой «соответствующих точек», были разработаны и стали достигли достоинства и определенности, хорошо поддающихся самоанализу, происходит ли вмешательство между реакциями двух (левого и правого) глазных систем.Достигнутое бинокулярное ощущение кажется объединенным из правого и левого монокулярных ощущений, выработанных независимо. (стр. 379)…. Таким образом, показано, что церебральные места зрительных образов для правого и левого глаза разделены. (стр. 380)
Как и в своей работе по рефлексам, Шеррингтон мыслит в терминах нейронных цепей, хотя и не определен:
Объединение ощущения составного источника, очевидно, связано с устройством нейронов, отличным от того, которое имеет место. в синтезе рефлекторного движения путем схождения рефлексов смежных дуг на его конечных общих путях.… Чистое соединение во времени без обязательного церебрального соединения в пространстве лежит в основе решения проблемы единства разума (стр. 381).
Шеррингтон завершает эту заключительную лекцию обсуждением произвольного контроля рефлексов, или, как он осторожно заявляет, «… рефлекторные дуги управляются механизмами, к активности которых добавляется сознание» (стр. 386). Он указывает на это как на механизм, с помощью которого мы приобретаем новые моторные навыки, что делает человека «… самым успешным животным на Земле в нынешнюю эпоху.'(Стр. 389). Его заключительное предложение по-прежнему верно:
Именно тогда вокруг головного мозга, его физиологических и психологических свойств должен в конечном итоге повернуться главный интерес биологии (стр. 390).
Удар
В 1957 году появилось несколько публикаций, посвященных столетию со дня рождения Шеррингтона. Среди них были следующие комментарии относительно воздействия The Integration Action на физиологические науки.
Лорд Адриан, получивший Нобелевскую премию 1932 года вместе с Шеррингтоном, прокомментировал:
Для неврологии было удачей, что его [Шеррингтона] пригласили в Йель в 1904 году для чтения лекций Силлимана. книга с названием Интегративное действие нервной системы , и именно в этой книге изложен план [обзор Шеррингтона управления моторикой в ответ на афферентные сигналы]. Это был новый подход к пониманию нервной системы, подход, который теперь настолько знаком, что мы принимаем его как должное во всех наших обсуждениях.(Адриан, 1957; стр. 214)
Сэр Джон Эклс, который начал свою научную карьеру с Шеррингтоном и позже (1963) лауреат Нобелевской премии, писал: надежные основы физиологии центральной нервной системы, и даже это Интегративное действие нервной системы оказало влияние, сравнимое с влиянием De motu cordis . … Исследования Шеррингтона нервной системы были задуманы в терминах анатомических концепций нейрона и синапса, и именно он показал, каким образом эти концепции важны для функционирования.Фактически, именно ясность его функционального мышления в терминах нейронов и синаптических связей между ними отличала Шеррингтона от всех современных нейрофизиологов. (Eccles, 1957; стр. 216 и 218)
Проф. Дерек Денни-Браун, который также начал свою карьеру с Шеррингтоном, прежде чем стать одним из выдающихся клинических неврологов своего времени, отметил:
Уже к 1906 году. … Шеррингтон почти в одиночку разгадал основные закономерности спинномозговых рефлексов и их взаимодействия.Это было колоссальное достижение, как и его ясное и классическое изложение в Integrative action , опубликованном в 1906 году, когда Шеррингтону было 49 лет. (Денни-Браун, 1957; стр. 544–545)
Постскриптум
Этот писатель мало что может или должен добавить к столь красноречивым оценкам. Но, возможно, мне разрешат личный комментарий. Я стал учеником Шеррингтона методом осмоса, когда учился в медицинской школе в конце 1950-х годов. К тому времени физиологи полностью усвоили подход Шеррингтона к нервной системе, заглянув за пределы экспериментальных данных в то, что они могут означать с точки зрения центральных механизмов.Я решил узнать больше о спинном мозге из-за встреч с пациентами, страдающими боковым амиотрофическим склерозом. В течение года исследования, выходящего из медицинской программы, я защитил диплом с отличием по супраспинальному контролю некоторых спинальных рефлексов под руководством моего наставника и друга, покойного доктора Уилбура К. Смита, клинического невролога и нейробиолога. Благодаря чтению проекта и связанных с ним клинических материалов моими научными героями стали сэр Джон Экклс и профессор Рагнар Гранит в области нейрофизиологии и доктор Дерек Денни-Браун в области клинической неврологии.Только позже, после того как я начал свою собственную работу в области нейрофизиологии в Национальном институте здоровья, я понял, что все трое начали свою научную карьеру с Шеррингтоном. К тому времени я купил переиздание The Integrative Action 1961 года и был интеллектуально готов понять его. Однако только со временем и многократным перечитыванием я смог полностью оценить его красоту и влияние. Для современных студентов будет откровением найти в нем так много основных понятий и терминологии, которые сейчас так глубоко укоренились в литературе, что стали их второй натурой.Я признателен редактору Brain за предоставленную мне возможность написать эту столетнюю дань уважения классическому научному труду и сэру Чарльзу Шеррингтону, который буквально произвел революцию в том, как мы все смотрим на мозг.
Список литературы
.Сэр Чарльз Скотт Шеррингтон, О. 1957–1952
,Примечания Rec R Soc Lond
,1957
, vol.12
(стр.211
—5
).Интеграционное действие Чарльза Скотта Шеррингтона: уведомление о столетнем юбилее
,JR Soc Med
,2004
, vol.97
(стр.34
—6
)Коэн Лорд Биркенхед
,Шеррингтон — физиолог, философ и поэт
,1958
Спрингфилд, Иллинойс
Чарльз К. Томас
.Школа физиологии Шеррингтона
,J Neurophysiol
,1957
, vol.20
(стр.543
—8
). ,Избранные труды сэра Чарльза Шеррингтона
,1979
Оксфорд, Великобритания
Oxford University Press
.Некоторые аспекты вклада Шеррингтона в нейрофизиологию
,Примечания Rec R Soc Lond
,1957
, vol.12
(стр.216
—25
).Жизнь в лаборатории Шеррингтона: его последнее десятилетие в Оксфорде 1925-1935
,Trends Neurosci
,1982
, vol.5
(стр.108
—10
),. ,Шеррингтон: его жизнь и мысли
,1979
Берлин
Springer International
.Die reflex Functionen des Zentralnerven mit besonderer Berucksichtigung, derhythmischen Tatigkeiten bein Sargeiter
,Ergenbnisse der Physiologie
,1916
, vol.15
(стр.480
—90
). ,Чарльз Скотт Шеррингтон: оценка
,1966
Лондон, Великобритания
Thomas Nelson and Sons, Ltd
.Чарльз Скотт Шеррингтон 1857-1952
,Извещения о некрологе Fellows R Soc
,1952
, vol.8
(стр.241
—70
). ,Открытие рефлексов
,1960
Лондон, Великобритания
Оксфорд: Clarendon Press
,.Рекрутмент и некоторые другие факторы подавления рефлексов
,Proc R Soc Lond Ser B
,1925
, vol.97
(стр.488
—518
). ,Histologie du système nerveux de l’homme et des vertébrés
,1909
Paris
Maloine
.,Гистология нервной системы человека и позвоночных
,1995
Нью-Йорк
Oxford University Press
.Заметки о расположении некоторых моторных волокон в пояснично-крестцовом сплетении
,J Physiol (Lond)
,1892
, vol.13
(стр.621
—772
).Дальнейшее экспериментальное замечание о корреляции действия мышц-антагонистов
,Proc Roy Soc
,1893
, vol.53
(стр.407
—20
).Замечание о коленном рефлексе и соотношении действия мышц-антагонистов
,Proc Roy Soc
,1893
, vol.52
(стр.556
—64
).Эксперименты по изучению периферического распределения волокон задних корешков некоторых спинномозговых нервов
,Philos Trans R Soc
,1894
, vol.184B
(стр.641
—763
).,Интегративное действие нервной системы
,1906
New Haven, CT
Yale University Press
. ,Интегративное действие нервной системы
,1947
New Haven, CT
Yale University Press
.Интеграция позы и движения: вклад Шеррингтона, Гесса и Бернстайна
,Hum Mov Sci
,2005
, vol.24
(стр.621
—43
),,,,,.,.Шеррингтон: гуманист, наставник и нейробиолог
,Classics in Movement Science
,2001
Champaign, IL
Human Kinetics
(стр.317
—74
).Интеграция рефлексов и двигателей
,Концепция интегративного действия Шеррингтона
,1969
Кембридж, Массачусетс, Массачусетс
Harvard University Press
© Автор (2007). Опубликовано Oxford University Press от имени Гарантов Мозга.Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
. .