2.2.2. Принципы координации рефлекторной деятельности
Взаимодействие возбуждающих и тормозных нервных процессов обеспечивает быстрое и точное приспособление организма к условиям существования. Такое взаимодействие процессов возбуждения и торможения в ЦНС, обеспечивающих ее согласованную деятельность, называют координацией.
Выделяют семь основных принципов координации рефлекторной деятельности:
конвергенция,
облегчение,
окклюзия,
принцип общего конечного пути,
принцип реципрокного торможения,
принцип обратной связи,
принцип доминанты.
Конвергенция — это морфологический принцип, который характеризуется схождением нескольких нейронов к меньшему их количеству.
Конвергенция, обеспечивающая схождение нервных импульсов к одному центральному нейрону, лежит в основе интегративной деятельности ЦНС. Она обусловлена схождением к телу и дендритам каждого нейрона аксонов множества других нервных клеток.
Схема организации конвергенции в ЦНС

1) вставочные нейроны, 2) эфферентный нейрон.
Конвергенция возбуждений лежит в основе облегчения рефлекторной реакции при одновременном раздражении нескольких афферентов.
Облегчение — это принцип координации рефлекторной деятельности, который характеризуется тем, что при одновременном раздражении двух афферентов возникает более сильная рефлекторная реакция, чем сумма рефлекторных ответов при раздельном возбуждении каждого из них.
Нервный центр состоит из двух зон: периферической и центральной. Центральная зона — это часть нервного центра, в которой из коллатералей афферента при передаче возбуждения выделяется достаточное количество медиатора для возникновения разрядной деятельности нейронов и рефлекторного ответа. При этом величина формирующегося ВПСП достаточна для того, чтобы ионные токи, циркулирующие между частично деполяризованной постсинаптической мембраной и аксонным холмиком, вызвали генерацию серии эфферентных ПД и рефлекторную реакцию.
Около 80% нейронов нервного центра локализуются в периферической зоне. Периферическая зона — это часть нервного центра, в которой из коллатералей афферента при передаче возбуждения выделяется недостаточное количество медиатора для возникновения разрядной деятельности нейронов и рефлекторного ответа. При этом величина ВПСП и циркулирующих местных ионных токов недостаточна для генерации в области аксонного холмика серии эфферентных ПД, а значит и рефлекторной реакции.
Облегчение рефлекторной реакции обусловлено перекрытием периферических зон расположенных рядом нервных центров. При одновременном раздражении двух афферентов, за счет конвергенции возбуждений в синапсах периферической зоны выделяется достаточное количество медиатора. Это обеспечивает возникновение разрядной деятельности нейронов, которые локализуются в периферической зоне, а значит усиление рефлекторного ответа.
Схема нейронной организации облегчения

1) афференты, 2) центральные зоны нервных центров, 3) периферические зоны нервных центров, 4) область перекрытия периферических зон.
Английский физиолог Ч. Шеррингтон установил, что одна и та же двигательная реакция может быть получена при раздражении различных афферентов. Это обусловлено тем, что отдельные мотонейроны являются общим конечным путем многочисленных сенсорных каналов. Принцип общего конечного пути («воронка» Шеррингтона) заключается в конкуренции множества афферентов и интернейронов за «обладание» одним общим конечным мотонейроном.
Схема нейронной организации общего конечного пути («воронка» Шеррингтона)

1) афференты, 2) вставочные нейроны, 3) мотонейрон.
Возбуждения различной сенсорной модальности и биологического качества конвергируют к одному мотонейрону. Из всего поступающего потока мотонейрон отбирает приоритетную (наиболее значимую) информацию и синтезирует эфферентную команду для исполнителей. При этом отбор приоритетной информации в ходе «борьбы» афферентов за «обладание» общим конечным мотонейроном обусловлен окклюзией возбуждений в нервных центрах.
Окклюзия — это принцип координации рефлексов, который характеризуется тем, что при одновременном раздражении двух афферентов сила рефлекторной реакции меньше, чем сумма рефлекторных ответов при раздельном возбуждении каждого афферента.
Окклюзия возникает вследствие перекрытия центральных зон нервных центров. При одновременном раздражении двух афферентов активируется меньше центральных нейронов, чем при раздельной их стимуляции, так как часть нервных клеток локализуется в области перекрытия центральных зон и являются общими для обоих нервных центров.
Схема нейронной организации окклюзии
1) афференты, 2) центральные зоны нервных центров, 3) область перекрытия центральных зон.
Принцип реципрокного (сопряженного) торможения характеризуется одновременным угнетением одного нервного центра при возбуждении другого.
Принцип реципрокности лежит в основе противоположных по функциональному назначению ответных реакций. Примером реципрокного торможения является взаимодействие нервных центров, обеспечивающих поочередное сокращение и расслабление мышц-сгибателей и разгибателей во время ходьбы, бега, плавания и т.п. Возбуждение, которое идет к мотонейронам, иннервирующим мышцы-сгибатели, по коллатерали афферента поступает к тормозным клеткам Реншоу. Эти клетки тормозят мотонейроны мышц-разгибателей. Если бы одновременно возбуждались нервные центры мышц сгибателей и разгибателей, движение конечности стало бы невозможным.
Схема нейронной организации реципрокного торможения жевательных мышц

1) собственно жевательная мышца, 2) мышца, опускающая нижнюю челюсть, 3) афферентные нейроны, 4) мотонейроны, 5) коллатераль, 6) тормозной нейрон.
Другим примером использования реципрокного торможения может быть стискивание зубов при сильной боли, чтобы подавить защитные движения, или прикусывание нижней губы, чтобы не смеяться при щекотании.
Нервные клетки обладают способностью регулировать частоту проходящих через них сигналов, а значит и силу рефлекторного ответа за счет обратной связи. Обратная связь базируется на способности коллатералей аксонов нервных клеток устанавливать синаптические контакты со вставочными нейронами, роль которых сводится к обратному воздействию на нейроны, образующие эти аксонные коллатерали.
Обратная связь может быть положительной и отрицательной. Положительная обратная связь осуществляется по принципу «чем больше, тем больше»: повышение частоты разрядной деятельности нейрона приводит к дальнейшему увеличению его возбудимости и импульсной активности. Механизм положительной обратной связи основан на реверберации возбуждения в замкнутых нейронных цепях, в которых имеются только возбуждающие нейроны.
Схема положительной обратной связи

1) эфферентный нейрон, 2) коллатераль, 3) возбуждающий вставочный нейрон.
Отрицательная обратная связь осуществляется по принципу «чем больше, тем» меньше: чем больше частота разрядной деятельности нейрона, тем меньше его возбудимость и импульсная активность. Механизм отрицательной обратной связи основан на реверберации возбуждения в нейронных ловушках, имеющих в своем составе тормозные нейроны.
Схема отрицательной обратной связи
1) эфферентный нейрон, 2) коллатераль, 3) тормозной вставочный нейрон.
Для деятельности нервной системы, как единого целого, в естественных условиях характерно наличие преобладающих (доминантных) очагов возбуждения, которые, по А.А. Ухтомскому (1923), являются проявлением одной из основных закономерностей работы нервных центров.
Доминанта — это господствующий очаг возбуждения в ЦНС, подчиняющий себе нервные центры в интересах выполнения наиболее важной в данный момент времени поведенческой деятельности организма.
Доминанту характеризуют пять основных свойств:
повышенная возбудимость,
способность к суммации,
стойкость возбуждения,
большая инертность,
способность реципрокно тормозить деятельность других нервных центров (тех, которые не связаны с организацией наиболее важной в данный момент поведенческой деятельности).
Доминанта усиливается за счет конвергенции афферентных возбуждений от различных рефлексогенных зон. Так, очаг доминантного возбуждения, который возникает при острой зубной боли, поддерживается посторонними раздражителями (прикосновение, яркий свет, резкий звук), которые усиливают страдания больного.
На способности доминантного очага угнетать другие нервные центры основан метод звуковой аналгезии (аудиоаналгезия). Звуковой рардражитель – звук или даже простой шум, создает в ЦНС доминанту, которая реципрокно тормозит очаги возбуждения, возникающие при различных стоматологических манипуляциях. При определенных условиях обезболивающий эффект аудиоаналгезии во время препарировании твердых тканей зуба может достигать 90%.
Принцип доминанты лежит в основе организации целенаправленной поведенческой деятельности, но не определяет ее адекватности биологическим и социальным потребностям человека. Примером этого могут служить алкогольная и наркотическая доминанты, которые, подчиняя себе большинство нервных центров с целью удовлетворения потребности в алкоголе или наркотическом веществе, разрушительно отражаются на физическом, психическом и социальном благополучии человека.
studfiles.net
19. Координационная деятельность цнс и ее структурно-функциональные основы.
В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности отдельных нервных клеток и нервных центров.
Координационная деятельность ЦНС – это согласованная деятельность различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними.
Основой координационной деятельности является взаимодействие процессов возбуждения и торможения.
Морфологическая основа координации: связь между нервными центрами (конвергенция, дивергенция, циркуляция).
20. Основные принципы координации: принцип реципрокности, окклюзии, суммации, иррадиации, принцип общего конечного пути, принцип обратной связи, принцип субординации, принцип доминанты.
1.Принцип реципрокности. Примером координационного взаимодействия рефлексов является реципрокная иннервация мышц-антагонистов.
2.Иррадиация означает способность к распространению нервного процесса (например, болевых ощущений) с места своего возникновения на другие участки центральной нервной системы. Она лежит в основе адаптации.
3.Общий конечный путь (принцип «воронки» Шеррингтона). Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути».
4.Обратная связь. Положительная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в том же направлении, что и основные сигналы, что ведет к усилению рассогласования в системе. Отрицательная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в противоположном направлении и направлены на ликвидацию рассогласования, т.е. отклонений параметров от заданной программы (П.К. Анохин).
5.Облегчение. Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов выше суммы реакций при изолированном раздражении этих рецептивных полей.
6.Окклюзия. Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов меньше, чем сумма реакций при изолированном раздражении каждого из рецептивных полей.
7. Доминанта. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в ЦНС. По А.А. Ухтомскому, доминантный очаг характеризуется:
— повышенной возбудимостью,
— стойкостью и инертностью возбуждения,
— способность к суммации возбуждений.
— тормозит другие очаги возбуждения
Принцип доминанты определяет формирование главенствующего возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
Доминанта есть общий принцип работы центральной нервной системы, она определяет освобождение организма от побочной деятельности во имя достижения наиболее важных для организма целей.
8. Субординация. Восходящие влияния преимущественно носят возбуждающий стимулирующий характер, нисходящие носят угнетающий тормозной характер. Эта схема согласуется с представлениями о росте в процессе эволюции роли и значении тормозных процессов в осуществлении сложных интегративных рефлекторных реакций. Имеет регулирующий характер.
studfiles.net
36. Основные принципы координационной деятельности цнс: реципрокности,
облегчения, окклюзии, обратной связи, общего «конечного» пути, доминанты.
Рефлекторная реакция в большинстве случаев осуществляется не одной, а целой группой рефлекторных дуг и нервных центров. Координация рефлекторной деятельности это такое взаимодействие нервных центров и проходящих по ним нервных импульсов, которое обеспечивает согласованную деятельность органов и систем организма. Она осуществляется с помощью следующих процессов:
1. Временное и пространственное облегчение. Это усиление рефлекторной реакции при действии ряда последовательных раздражителей или одновременном их воздействии на несколько рецептивных полей. Объясняется явлением суммации в нервных центрах.
2. Окклюзия явление противоположное облегчению. Когда рефлекторная реакция на два или более сверхпороговых раздражителя меньше, чем ответы на их раздельное воздействие. Оно связано с конвергенцией нескольких возбуждающих импульсов на одном нейроне.
3. Принцип общего конечного пути. Разработан Ч. Шеррингтоном. В основе его лежит явление конвергенции. Согласно этому принципу на одном эфферентном мотонейроне могут образовывать синапсы нескольких афферентных, входящих в несколько рефлекторных дуг. Этот нейрон называется общим конечным путем и участвует в нескольких рефлекторных реакциях. Если взаимодействие этих рефлексов приводит к усилению общей рефлекторной реакции, такие рефлексы называются союзными. Если же между афферентными сигналами происходит борьба за мотонейрон — конечный путь, то антагонистическими. В результате этой борьбы второстепенные рефлексы ослабляются, а жизненно важным освобождается общий конечный путь.
4. Реципрокное торможение. Обнаружено Ч. Шеррингтоном. Это явление
торможения одного центра в результате возбуждения другого. Т.е. в этом случае тормозится антагонистический центр. Например при возбуждении центров сгибания левой ноги, по реципрокному механизму тормозятся центры мышц разгибателей этой же ноги и центры сгибателей правой. В реципрокных взаимоотношениях находятся центры вдоха и выдоха продолговатого мозга, центры сна и бодрствования и т.д.
5. Принцип доминанты. Открыт А.А. Ухтомским. Доминанта — это преобладающий очаг возбуждения в ЦНС, подчиняющий себе другие НЦ. Доминантный центр обеспечивает комплекс рефлексов, которые необходимы в данный момент для достижения определенной цели. При некоторых условиях возникают питьевая, пищевая, оборонительная, половая и др. доминанты. Свойствами доминантного очага являются повышенная возбудимость, стойкость возбуждения, высокая способность к суммации, инертность. Эти свойства обусловлены явлениями облегчения, иррадиации, с одновременным повышением активности вставочных тормозных нейронов, которые тормозят нейроны других центров.
6. Принцип обратной афферентации. Результаты рефлекторного акта воспринимаются нейронами обратной афферентации и информация от них поступает обратно в нервный центр. Там они сравниваются с параметрами возбуждения и рефлекторная реакция корректируется.
studfiles.net
Основные принципы координационной деятельности цнс
В основе координационной деятельности ЦНС лежит взаимодействие между процессами возбуждения и торможения. О существовании возбуждения в нервах, мышцах, в ЦНС было известно давно. Торможение в ЦНС было открыто И.М.Сеченовым (1862 г.) в опытах на лягушках и получило название «Сеченовское торможение». Он определял время сгибательного рефлекса (по Тюрку), погружая лапку лягушки в кислоту, а затем на зрительные бугры накладывал кристаллик поваренной соли. После наложения кристаллика происходило удлинение времени рефлекса или рефлекс полностью затормаживался, а после снятия кристаллика соли и промывания этого участка мозга водой время рефлекса восстанавливалось до исходного уровня. Согласованная (координационная) деятельность обеспечивается за счет ряда механизмов:
1) Принцип доминанты. Он был сформулирован А.А.Ухтомским как основной принцип работы нервных центров. Доминантный (или господствующий) очаг возбуждения характеризуется следующими свойствами: повышенной возбудимостью; инертностью (стойкостью) возбуждения, т.е. может сохраняться длительное время; способностью к суммации возбуждений, притягивая на себя возбуждение с других центров; способностью тормозить субдоминантные очаги возбуждения других нервных центров.
2) Принцип окклюзии. Этот принцип противоположен пространственному облегчению или суммации, и он заключается в том, что два афферентных входа совместно возбуждают меньшую группу мотонейронов по сравнению с эффектами при раздельной их активации. Причина окклюзии состоит в том, что афферентные входы в силу конвергенции отчасти адресуются к одним и тем же мотонейронам, которые затормаживаются при активации обоих входов одновременно. Явление окклюзии проявляется в случаях применения сильных афферентных раздражений.
3) Принцип обратной связи. Процессы саморегуляции в организме в полном объеме могут осуществляться только в том случае, когда функционирует канал обратной связи. За счет импульсов, поступающих по этому каналу, происходит оценка правильности исполнения поставленной задачи, а если она не выполнена, то вносятся коррекции для достижения результата.
Велико значение механизмов обратной связи в поддержании гомеостаза. Так, например, поддержание постоянного уровня кровяного давления осуществляется за счет изменения импульсной активности барорецепторов сосудистых рефлексогенных зон, в результате чего измененяеся тонус вазомоторных симпатических нервов и таким образом нормализуют кровяное давление.
4) Принцип реципрокности (сочетанности, сопряженности, взаимообусловленности). Он отражает характер отношений между центрами ответственными за осуществление противоположных функций (вдоха и глотания, выдоха и выдоха, сгибания и разгибания конечностей и т.д.). Например, активация проприорецепторов мышцы-сгибателя одновременно возбуждает центр мышц сгибателей и тормозит центр мышц разгибателей. Реципрокное торможение играет важную роль в координации двигательных актов. Реципркные отношения имеют динамический характер (о чем говорил еще Введенский), а Шеррингтон эти отношения рассматривал как статические явления. Опытами П.К.Анохина с перекрестным подшиванием сухожилий сгибателей к разгибателям и наоборот было установлено, что через 6–8 месяцев мышцы сгибатели начинают выполнять функцию разгибателей, а разгибатели функцию сгибателей. Такая перестройка реципрокных взаимоотношений была бы невозможна, если реципрокные отношения имели бы строго раз и навсегда зафиксированный (статический) характер. За счет пластичности ЦНС и в результате постоянной неадекватной импульсации с сокращающихся мышц происходит изменение первоначального функционального взаимоотношения между сгибательным и разгибательным центрами. Эти исследования Анохина, проведенные еще в 30–е годы, послужили основой для введения понятия обратная афферентация (шестой составной части рефлекторного пути) и явились основой для создания теории о функциональных системах и биологической кибернетики (опередив в этом отношении Винера, который считается основоположником кибернетики (1948 г.), примерно на 13–15 лет).
5) Принцип общего конечного пути. Эффекторные нейроны ЦНС, например, мотонейроны спинного мозга, могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого числа афферентных и промежуточных нейронов, для которых они являются конечным путем (путем от ЦНС к эффектору). Например, на мотонейронах передних рогов спинного мозга, иннервирующих мускулатуру конечности, оканчиваются волокна афферентных нейронов, нейронов пирамидного тракта и экстрапирамидной системы (ядер мозжечка, ретикулярной формации и многих других структур)
6) Явления конвергенции — схождение нервных импульсов на одни и те же центральные нейроны. Такая особенность зависит не только от функциональных свойств центров, но обусловлено также количественными соотношениями периферических рецепторных и промежуточных центральных нейронов. Это соотношение составляет примерно 10:1. Явления конвергенции играют решающее значение при формировании общего конечного пути.
7) Явления дивергенции — процесс противоположный конвергенции, т.е. импульсы, поступающие в ЦНС, распространяются (иррадиируют) на соседние участки.
8) Субординационные взаимоотношения — соподчинение, т.е. вышележащие отделы ЦНС оказывают свое регулирующее влияние на нижележащие отделы.
studfiles.net
Координация функций в ЦНС | Кинезиолог
Принципы координации в ЦНС
Координация – это согласование и сопряжение нервных процессов, характерное для деятельности центральной нервной системы (ЦНС).
1. Принцип реципрокной (сопряженной, взаимоисключающей) иннервации.
2. Принцип общего конечного пути (принцип конвергенции, «воронка Ч. Шеррингтона»).
3. Принцип доминанты.
4. Принцип временнОй связи.
5. Принцип саморегуляции (прямая и обратная связи).
6. Принцип иерархии (соподчинение).
1. Принцип реципрокной (сопряженной, взаимоисключающей) иннервации
Принцип реципропной иннервации мышц-антагонистов впервые был обнаружен в 1896 году выдающимся отечественным физиологом Н.Е. Введенским, учеником И.М. Сеченова.
Сокращение сгибателя вызывает понижение тонуса разгибателя на той же стороне, а с противоположной
стороны — наоборот: может вызывать повышение тонуса разгибателя.
На реципрокном принципе основан рефлекс шагания. Таким образом, ходьба — это условнорефлекторная, основанная на принципе реципрокной иннервации, циклическая двигательная деятельность ног.
Вывод:
Возбуждение сгибателя вызывает сопряженное торможение и расслабление разгибателя: происходит перекрестный разгибательный рефлекс.
2. Принцип общего конечного пути (принцип конвергенции)
Этот принцип был открыт и исследован выдающимся английским физиологом сэром Ч.С. Шеррингтоном (Charles Scott Sherrington) в 1896 году.
Он установил, что в нервных центрах количество афферентных (приносящих) клеток намного больше, чем количество эфферентных (выносящих) нейронов, несущих возбуждение к мышцам. Получается, что между нейронами идет борьба «за общий конечный путь», т.е. за то, чтобы передать своё возбуждение на эффентные нейроны. Этот принцип получил также образное наименование «воронки Шеррингтона».
3. Принцип доминанты
Доминанта (от лат. «господствовать») — это временно господствующий рефлекс, подчиняющий себе дуги прочих рефлексов. Доминанта существует в виде устойчивого очага возбуждения, подчиняющего себе другие возбуждённые очаги.
Доминанта может быть гуморальной, или её можно вызвать искусственно, если вызвать деполяризацию участка головного мозга с помощью химического или электрического воздействия.
Примеры доминанты:
Попытки лягушки снять себя с крючка.
Особенности доминантного очага (центра) :
— повышенная возбудимость,
— повышенная стойкость (устойчивость к тормозным воздействиям),
— тормозное воздействие на другие возбуждающиеся очаги,
— способность к суммации возбуждения с соседних участков,
— длительность существования данного возбуждённого очага,
— инерция, т.е. длительное удержание возбуждённого состояния после прекращения первоначального возбуждения и сопротивление тормозному воздействию.
Доминанта была открыта в 1924 году А.А. Ухтомским, крупным отечественным физиологом, учеником другого крупного физиологи — Н.Е. Введенского.
Суть этого явления заключается в том, что если существует доминантный очаг, имеющий возбуждение, то любое другое возбуждение будет усиливать реакцию именно этого доминантного очага. И рефлекторный ответ будет соответствовать именно доминантному очагу (доминантному нервному центру), а не раздражителю. Можно сказать, что доминанта нарушает протекание классических условных и безусловных рефлексов. Кроме того, доминантный очаг тормозит все другие центры и подавляет их возбуждение. Таким образом, доминанта как бы фильтрует возбуждение, приходящее из разных источников, т.к. тормозит все посторонние ненужные импульсы.
В 1960-е годы В.С. Русинов получил искусственную доминанту путем слабого электрического раздражения 6-го слоя коры больших полушарий головного мозга.
Иногда в основе доминанты лежит снижение лабильности (подвижности нервных процессов).
Формы доминанты
1. Чувствительная (сенсорная).
2. Двигательная.
По механизму:
1. Рефлекторная.
2. Гуморальная (голодовая, половая).
По уровню расположения:
1. Спинальная (спинной мозг).
2. Бульбарная (продолговатый мозг).
3. Мезэнцефальная (средний мозг).
4. Диэнцефальная (промежуточный мозг).
5. Кортикальная (корковая).
4. Принцип временной связи
Высшая форма временной связи — условный рефлекс.
5. Принцип саморегуляции (прямая и обратная связи)
Прямые и обратные связи — это пути влияния управляющего объекта на управляемый объект. Соответственно, влияние может быть прямым и обратным.
Обратные связи, в свою очередь, делятся на положительные (усиливающие) и отрицательные (ослабляющие).
6. Принцип иерархии (соподчинение)
Принцип иерархии очень прост — нижележащие структуры подчиняются вышележащим. Это означает, что вышележащие структуры умеют как подгонять, так и тормозить нижележащие структуры.
Существует также функциональная иерархия. Так, высшее место в иерархии безусловных рефлексов занимает оборонительный рефлекс, затем — пищевой рефлекс, потом половой. Но в ряще случаев лидерство может захватывать половой рефлекс, оттесняя на второй план пищевое поведение и даже инстинкт самосохранения.
Дополнительные материалы
Принципы координационной деятельности мозга
kineziolog.su
Торможение в ЦНС. Принципы координационной деятельности ЦНС
Министерство здравоохранения Республики Беларусь
УО «Гомельский государственный медицинский университет»
Кафедра нормальной физиологии
Обсуждено на заседании кафедры
Протокол №__________200__года
ЛЕКЦИЯ №25.
по нормальной физиологии для студентов 2 курса
Тема: Торможение в ЦНС. Принципы координационной деятельности ЦНС.
Время 90 минут
Учебные и воспитательные цели:
Изучить природу, функциональное значение, структурно-функциональную организацию торможения в центральной нервной системе.
ЛИТЕРАТУРА
Физиология человека. Под ред. В.М.Покровского, Г.Ф.Коротько.М., Медицина. – 2000.-Т..1 — С 115 – 126.
Мультимедийная презентация 34 слайда.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
№ п/п | Перечень учебных вопросов | Количество выделяемого времени в минутах |
1. | Введение | 5 |
2 | Торможение в ЦНС. Природа торможения. Виды торможения. | 25 |
3 | Первичное постсинаптическое торможение | 25 |
4 | Пресинаптическое торможение | 10 |
5 | Вторичное торможение | 10 |
6 | Механизмы координации рефлекторной деятельности | 5 |
7 | Компенсация функций и пластичность нервных центров | 5 |
8 | Заключение | 5 |
Всего 90 мин
Торможение в ЦНС. Природа торможения. Виды торможения. Взаимосвязь возбуждения и торможения в ЦНС.
Торможение в ЦНС — особый нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения.
Классификация и механизмы центрального торможения:
1. Первичное постсинаптическое торможение — торможение , не- связанное с первоначальным процессом возбуждения и развивающееся в результате активации специальных тормозных структур. Тормозные синапсы образуют в своих окончаниях тормозной медиатор (ГАМК, глицин, в отдельных синапсах ЦНС роль тормозного медиатора может играть ацетилхолин). На постсинаптической мембране развивается тормозной постсинаптический потенциал (ТПСП), снижающий возбудимость мембраны постсинаптического нейрона. Тормозными нейронами могут служить только вставочные нейроны, афферентные нейроны всегда являются возбуждающими. В зависимости от вида тормозных нейронов и структурной организации нейронной сети постсинаптическое торможение подразделяется на:
Реципрокное торможение. Оно лежит в основе функционирования мышц-антагонистов и обеспечивает расслабление мышцы в момент сокращения мышцы-антагониста. Афферентное волокно, проводящее возбуждение от проприорецепторов мышц (например, сгибателей), в спинном мозге делится на две ветви: одна из них образует синапс на мотонейроне, иннервирующем мышцу-сгибатель, а другая — на вставочном, тормозном, образующем тормозной синапс на мотонейроне, иннервирующем мышцу-разгибатель. В результате возбуждение, приходящее по афферентному волокну, вызывает возбуждение мотонейрона, иннервирующего сгибатель и торможение мотонейрона мышцы-разгибателя.
Возвратное торможение. Оно реализуется через тормозные клетки Реншоу, открытые в спинном мозге. Аксоны мотонейронов передних рогов отдают коллатераль на тормозной нейрон Реншоу, аксоны которого возвращаются на тот же мотонейрон, образуя на нем тормозные синапсы. Таким образом формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов мотонейрона.
Центральное (Сеченовское) торможение. Оно осуществляется тормозными вставочными нейронами, через которые реализуется влияние на мотонейрон спинного мозга возбуждения, возникающего в зрительных буграх под влиянием их раздражения. На мотонейроне спинного мозга суммируются ВПСП, возникающие в болевых рецепторах конечности и ТПСП, возникающие в тормозных нейронах под влиянием возбуждения таламуса и ретикулярной формации. В результате время защитного сгибательного рефлекса возрастает.
Латеральное торможение осуществляется с помощью тормозных вставочных нейронов в параллельных нейронных сетях.
Первичное пресинаптическое торможение развивается в терминальных отделах аксонов (перед пресинаптической структурой) под влиянием специальных аксо-аксональных тормозных синапсов. Медиатор этих синапсов вызывает деполяризацию мембраны терминалей и приводит их в состояние, подобное катодической депрессии Вериго. Мембрана в области такого бокового синапса препятствует проведению потенциалов действия к пресинаптической мембране, активность синапса уменьшается.
Пресинаптическое торможение
Пресинаптическим торможением называют снижение или выключение активности клетки за счет синаптического торможения оканчивающейся на ней возбуждающей терминали. Явление пресинаптического торможения зафиксировали Гассер и Грэхем в 1933 г., наблюдая эффект развития торможения сгибательных рефлексов при раздражении других корешков. Данный вид торможения термином “пресинаптическое торможение” впервые обозначили Фрэнк и Фуортес в 1957 г.
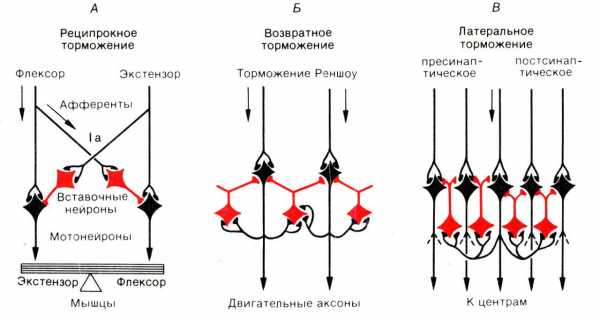
Рис. 1. Электронная микрофотография пресинаптических тормозных синапсов. Обозначения: пре. и пост. – пре- и постсинаптический контакт нейрона.
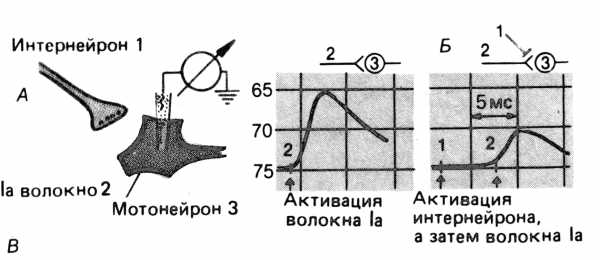
Рис. 2. Пресинаптическое торможение.
А. Схема эксперимента для демонстрации пресинаптического торможения моносинаптических ВПСП в мотонейроне.
Б. ВПСП при стимуляции гомонимных волокон Ia без (слева) и после (справа) предварительной активации пресинаптических тормозных интернейронов.
По “Физиология человека” под ред Р.Шмидта. – 1996. – Т.1. – С.59.
Стимуляция тормозного интернейрона подавляет моносинап-тический ВПСП мотонейрона.
Рис. 3. Подавление моносинаптического ВПСП при синаптическом торможении.
По Дж. Экклс. Физиология синапсов. – М.: Мир. – 1966. – С279.
Развитие этого подавления ВПСП протекает с латентным периодом, равным 5 мс, максимум достигается через 20 мс, а общая длительность превосходит 200 мс.
Рис. 4. Временной ход пресинаптического торможения моносинаптических возбуждающих ВПСП.
По “Физиология человека” под ред Р.Шмидта. – 1996. – Т.1. – С.59.
Увеличение частоты предварительных раздражений изменяет характер подавления. В частности, одна серия стимуляции с частотой 200-300 импульсов в секунду вызывает максимальное подавление менее чем на 10%, а две серии – подавление менее чем на 20%. При пресинаптическом торможении подавление моносинаптического ВПСП не связано с какими-либо изменениями их временных параметров.
Тормозные синапсы на окончаниях волокон Ia обеспечивают довольно значительную деполяризацию, называемую деполяризацией первичных афферентов, или же первичную эфферентную деполяризацию (ПАД). В спинном мозгу ПАД обнаруживает длительную фазу (до 25 мс) нарастания до закругленной вершины и характеризуется большей продолжительностью по сравнению с постсинаптическими процессами. Большая продолжительнсоть ПАД объясняется или длительным действием медиатора, или медленным, пассивным снижением деполяризации вследствие большой электрической постоянной времени мембраны. Пассивно снижающаяся компонента ПАД снимается импульсом, распространяющимся по афферентному волокну до его центральных окончаний.
Существует соответствие во всех отношениях между наблюдаемой деполяризацией первичных афферентных волокон и подавлением их синаптического возбуждающего действия.
Пресинаптическая деполяризация афферентов уменьшает величину их пресинаптического спайкового потенциала и таким образом уменьшает вызываемый им ВПСП. По данным Каца (1962), снижение спайкового потенциала на 5 мВ приводит к снижению выброса квантов медиатора и к снижению ВПСП до 50% и менее.
Характер ПАД в различных нейронах отличается по своим характеристикам. В целом временные параметры сравнимы. ПАД волокон кожного нерва отличается большей величиной амплитуды на одиночные раздражения с более коротким латентным периодом (около 2 мс), максимум также достигается ранее, чем в случае ПАД, вызываемых ритмическим раздражением нервных волокон, идущих от мышц. ПАД в клиновидном ядре имеет короткий латентный период (около 2 мс) и быстрый подъем до максимума.
Тормозные синапсы имеют химическую природу, медиатором в них служит ГАМК. Деполяризация первичных афферентов инактивирует возбуждающие натриевые каналы.Шунтирование натриевых каналов снижает амплитуду пресинаптического потенциала действия. В результате синаптическая передача моторного импульса ослабляется или исключается.
Во всех типах возбуждающих синапсов обнаруживается тесная зависимость между деполяризацией пресинаптических волокон и торможением синаптической передачи. Пресинаптическое торможение оказывает влияние на возбуждающие синапсы всех типов, образуемые афферентными волокнами, входящими в спинной мозг, групп Ia, Ib и II, отходящими от мышц, и альфа-волокнами кожного нерва. Это торможение влияет не только на местные спинномозговые рефлексы, но также и на синаптические передачи в восходящих путях как от кожных афферентов, так и на спиноцеребеллярные. Кроме того, пресинаптическое торможение влияет на синаптические передачи задних столбов в ядра нежного и клиновидного пучков. Нисходящие импульсы из коры головного мозга и ствола мозга также оказывают пресинаптическое тормозное влияние на волокна группы I и кожные афферентные волокна в спинном мозгу и клиновидном ядре. Обнаружено пресинаптическое торможение вторичных афферентных волокон отходящих от клиновидного ядра и имеющих переключение в таламусе. Синапсы с пресинаптическим торможением обнаружены в связанном с таламусом ядре мозга – латеральном коленчатом теле. В коре головного мозга не выявлено синаптических структур, которые могли бы осуществлять пресинаптическое торможение. На этих высших уровнях нервной системы доминирует постсинаптическое торможение. Пресинаптическое торможение действует как отрицательная обратная связь, уменьшая приток сенсорной информации в центральную нервную систему. Обычно эта отрицательная обратная связь не имеет точной топографии, но обычно концентрируется в пределах одной сенсорной модальности.
Пресинаптическое торможение служит механизмом регуляции двигательных систем спинного мозга. Его особенностью является возможность специфического воздействия на отдельные синаптические входы без изменений возбудимости всей клетки. Таким образом, избыточная информация устраняется еще до того, как достигает места интеграции клеточного тела нейрона.
2. Вторичное торможение не связано с тормозными структурами, является следствием предшествующего возбуждения. Пессимальное торможение (открыто Н.Е.Введенским в 1886 г.) развивается в полисинаптических рефлекторных дугах при чрезмерной активации центральных нейронов и играет предохранительную роль. Оно выражается в стойкой деполяризации мембраны, приводящей к инактивации натриевых каналов. «Торможение вслед за возбуждением» развивается в нейронах непосредственно после потенциала действия и характерно для клеток с длительной следовой гиперполяризацией. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов активности нейронов.
Механизмы координации рефлекторной деятельности: реципрокная иннервация, доминанта (А.А.Ухтомский), принципы обратной связи и общего конечного пути, принцип субординации.
Принцип иррадиации возбуждения. Иррадиация — распространение, расширение рефлекторного ответа. Это феномен “растекания” возбуждения по нейронам центральной нервной системы, развивающийся или после действия сверхсильного раздражителя, или на фоне выключения торможения. Распространение возбуждения возможно за счет многочисленных контактов между нейронами, возникающих при ветвлении аксонов и дендритов вставочных нейронов. Иррадиация позволяет увеличивать количество участвующих в рефлекторном ответе групп мышц. Ограничивают иррадиацию тормозные нейроны и синапсы.
На фоне действия стрихнина, блокирующего тормозные синапсы, наступают генерализованные судороги при тактильной стимуляции любого участка тела или при раздражении рецепторов любой сенсорной системы. В коре больших полушарий наблюдается явление иррадиации процесса торможения.
В основе координации рефлекторных актов лежат определенные механизмы, основанные на структурно-функциональной организации ЦНС и обозначаемые как “принципы” формирования рефлекторного ответа.
Принцип реципрокной иннервации. Реципрокная (сопряженная) координация открыта Н.Е.Введенским в 1896 году. Обусловлена реципрокным торможением, т.е. активация одного рефлекса одновременно сопровождается торможением второго, противоположного по своей физиологической сущности.
Принцип общего «конечного пути«. Открыт английским физиологом Ч.Шеррингтоном (1906). Один и тот же рефлекс (например, сокращение мышцы) может быть вызван раздражением различных рецепторов, т.к. один и тот же конечный -мотонейрон передних рогов спинного мозга входит в состав многих рефлекторных дуг. Рефлексы, дуги которых имеют общий конечный путь, подразделяются на агонистические и антагонистические. Первые усиливают, вторые тормозят друг друга, как бы конкурируя за конечный результат. В основе подкрепления лежит конвергенция и суммация, в основе конкуренции за конечный путь — сопряженное торможение.
Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром. Обратная связь состоит во вторичной афферентации, поступающей в ЦНС от рецепторов, которые возбуждаются при изменении функциональной активности рабочего органа. Например, потенциалы действия, обусловленные возбуждением рецепторов мышц, сухожилий и суставных сумок сгибающейся конечности, в процессе осуществления акта сгибания поступают во все структуры ЦНС, начиная от центров спинного мозга. Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и отрицательную, когда рефлекс, ее вызывающий, тормозится. Обратная связь лежит в основе саморегуляции функций организма.
Принцип отдачи. Феномен отдачи состоит в быстрой смене одного рефлекса другим противоположного значения. Например, после сгибания конечности ее разгибание происходит быстрее, особенно если сгибание было сильным. Механизм этого явления состоит в том, что при сильном сокращении мышц возбуждаются рецепторы Гольджи сухожилий, которые через тормозные вставочные нейроны тормозят мотонейроны сгибательных мышц и образуют ветвь, которая возбуждает центр мышц — разгибателей. Благодаря этому механизму можно получить сумму рефлексов — цепные рефлексы (окончание одного рефлекторного ответа инициирует следующий) и ритмические (многократное повторение ритмичных движений).
Принцип доминанты. Конечный поведенческий эффект при координации рефлексов может быть изменен в зависимости от функционального состояния центров (наличия доминантных очагов возбуждения).
Особенности доминантного очага возбуждения:
1. Повышенная возбудимость нейронов.
2. Стойкость процесса возбуждения.
3. Способность к суммации возбуждения.
4. Иннертность. Очаг доминирует, подавляет соседние центры путем сопряженного торможения, возбуждаясь за их счет. Доминанту можно получить химическим воздействием на центры, например, стрихнином. В основе доминантного возбуждения лежит способность возбудительного процесса к иррадиации по нейронным цепям.
Компенсация функций и пластичность нервных центров
Компенсаторная функция нервных центров заключается в том, что даже после значительных разрушений части нервных клеток и глиальных элементов происходит восстановление функций центра. В отдельных случаях центр способен перестроиться таким образом, что может принимать участие в регуляции совершенно иной функции.
Пластичность — это способность нервных центров изменять свое прямое функциональное назначение и расширять свои функциональные возможности. Это качество генетически запрограммировано, но может значительно развиваться под влиянием упражнений. Пластичность ЦНС проявляется при травмах, когда функция поврежденного участка (например, коры больших полушарий головного мозга) обеспечивается за счет сохранившихся или смежных отделов.
Заведующий кафедрой, профессор Э.С. Питкевич
Дата переработки 8 декабря 2006г.
7
studfiles.net
(реципрокное, возвратное, латеральное), их значение. Тпсп.
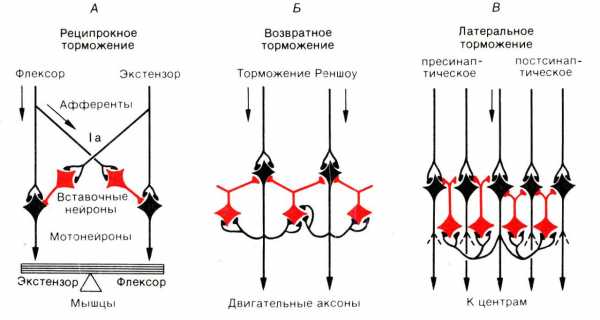
Реципрокное (сопряженное)
Возвратное
Латеральное
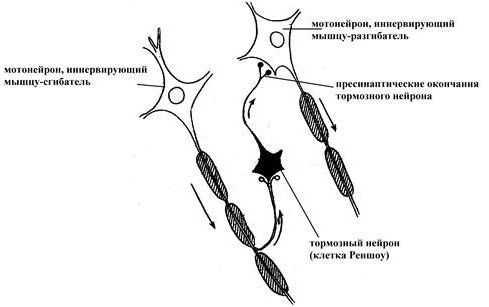
Координированная работа антагонистических нервных центров обеспечивается формированием реципрокных отношений между нервными центрами благодаря наличию специальных тормозных нейронов – клеток Реншоу.
Известно, что сгибание и разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Сигнал от афферентного звена через промежуточный нейрон вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель, а через клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель (и наоборот)
Угнетение активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки с участием тормозного вставочного нейрона. Например, мотонейрон переднего рога спинного мозга дает боковую коллатераль, которая возвращается назад и заканчивается на тормозных нейронах – клетках Реншоу. Аксон клетки Реншоу заканчивается на том же мотонейроне, оказывая на него тормозное действие (принцип обратной связи).
Возбуждение, передаваемое через коллатерали аксона возбужденной нервной клетки, активирует вставочные тормозные нейроны, которые тормозят активность соседних нейронов, в которых возбуждение отсутствует или является более слабым.
В результате в этих соседних клетках развивается очень глубокое торможение. Образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону.
Латеральное торможение по нейронному механизму действия может иметь форму как постсинаптического, так и пресинаптического торможения. Играет важную роль при выделении признака в сенсорных системах, коре больших полушарий.
18. Роль торможения в адаптивной деятельности организма.
1 — Координация рефлекторных актов. Направляет возбуждение к определенным нервным центрам или по определенному пути, выключая те нейроны и пути, деятельность которых в данный момент является несущественной. Результатом такой координации является определенная приспособительная реакция.
2 -Ограничивают иррадиацию возбуждения и концентрируют его в определенных отделах ЦНС;
3 — Охранительное. Предохраняет нервные клетки от перевозбуждения и истощения. Особенно при действии сверхсильных и длительно действующих раздражителей.
19. Координационная деятельность цнс и ее структурно-функциональные основы.
В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности отдельных нервных клеток и нервных центров.
Координационная деятельность ЦНС – это согласованная деятельность различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними.
Основой координационной деятельности является взаимодействие процессов возбуждения и торможения.
Морфологическая основа координации: связь между нервными центрами (конвергенция, дивергенция, циркуляция).
20. Основные принципы координации: принцип реципрокности, окклюзии, суммации, иррадиации, принцип общего конечного пути, принцип обратной связи, принцип субординации, принцип доминанты.
1.Принцип реципрокности. Примером координационного взаимодействия рефлексов является реципрокная иннервация мышц-антагонистов.
2.Иррадиация означает способность к распространению нервного процесса (например, болевых ощущений) с места своего возникновения на другие участки центральной нервной системы. Она лежит в основе адаптации.
3.Общий конечный путь (принцип «воронки» Шеррингтона). Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути».
4.Обратная связь. Положительная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в том же направлении, что и основные сигналы, что ведет к усилению рассогласования в системе. Отрицательная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в противоположном направлении и направлены на ликвидацию рассогласования, т.е. отклонений параметров от заданной программы (П.К. Анохин).
5.Облегчение. Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов выше суммы реакций при изолированном раздражении этих рецептивных полей.
6.Окклюзия. Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов меньше, чем сумма реакций при изолированном раздражении каждого из рецептивных полей.
7. Доминанта. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в ЦНС. По А.А. Ухтомскому, доминантный очаг характеризуется:
— повышенной возбудимостью,
— стойкостью и инертностью возбуждения,
— способность к суммации возбуждений.
— тормозит другие очаги возбуждения
Принцип доминанты определяет формирование главенствующего возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
Доминанта есть общий принцип работы центральной нервной системы, она определяет освобождение организма от побочной деятельности во имя достижения наиболее важных для организма целей.
8. Субординация. Восходящие влияния преимущественно носят возбуждающий стимулирующий характер, нисходящие носят угнетающий тормозной характер. Эта схема согласуется с представлениями о росте в процессе эволюции роли и значении тормозных процессов в осуществлении сложных интегративных рефлекторных реакций. Имеет регулирующий характер.
studfiles.net
